Stromy sú komplexné prírodné objekty,
pozostávajúce z mnohých opakujúcich sa (podobných) častí. Na ich
modelovanie, podobne ako aj na modelovanie iných prírodných objektov a javov,
sa v počítačovej grafike používajú procedurálne metódy, ktoré
umožňujú na základe relatívne malého počtu vstupných parametrov
generovať na pohľad veľmi zložité objekty, a teda aj stromy (ako
bohato rozkonárené štruktúry).
V práci prinášame
prehľad problematiky súvisiacej s procedurálnym modelovaním stromov.
Pre lepšie pochopenie biologickej podstaty stromov sme v druhej kapitole
uviedli prehľad základných botanických pravidiel uplatňujúcich sa pri
raste a rozkonárovaní stromov. V tretej kapitole hovoríme
o procedurálnych technikách, medzi ktoré patria aj L-systémy
a algoritmy na generovanie fraktálov. Fraktálne vlastnosti rastlín
popisujeme v štvrtej kapitole a teóriu L-systémov v piatej
kapitole. V šiestej kapitole sa venujeme geometrickej reprezentácii
stromov, pričom dôkladnejšie popisujeme rôzne detailné reprezentácie ich
tvaru a štruktúr, so zameraním sa na modulárnu štruktúru rastlín. Siedma
kapitola konečne prináša prehľad hlavne procedurálnych, ale aj
interaktívnych techník použitých na modelovanie stromov. V ôsmej kapitole
je prehľad možného využitia virtuálnych modelov rastlín.
Pri modelovaní stromov (drevín) je veľmi potrebné
poznať ich biologickú stavbu a funkciu. Nasledujúca botanická
časť je spracovaná podľa (Vĕtvička et al., 1992, Bobák
et al., 1992)
Dreviny
sú rastliny celé drevnaté, drevnatejú im aj tohtoročné výhonky, ktoré
v nasledujúcom roku pokračujú v raste. Sú viacročné až
dlhoveké, plodia veľa ráz a ich stonky sa obyčajne rozkonárujú.
Podľa vzhľadu sa rozdeľujú na stromy, drevnaté liany a kry.
Stromy
sú dreviny so stonkou (nadzemný orgán, ktorý spája orgány výživy – korene a
listy) rozkonárenou až v určitej výške nad zemou, takže majú dve
odlišné časti, nerozkonárený kmeň,
ktorý prechádza do bohato rozkonárenej koruny.
Napriek tomu, že stonka je najtvárnejší
a najpremenlivejší orgán rastlinného tela, riadi sa výstavba každej koruny
dreviny presnými dedičnými zákonitosťami, hoci vonkajšie ekologické
vplyvy alebo patologické stavy môžu spôsobiť vážne zmeny vo vzhľade
jedinca.
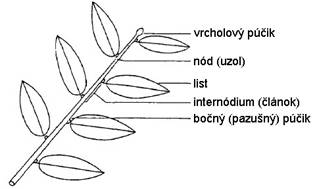
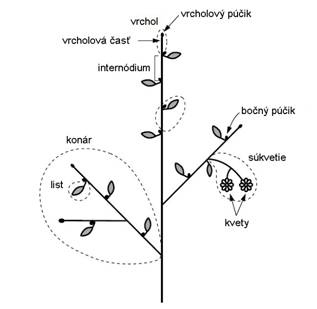
Stonka
je článkovaná, rozdelená na uzly (nódy)
a články (internódiá) (Obr. 2.1). Článkovanosť stonky súvisí so vznikom a
pripojením listov a bočných stoniek. Uzol je miesto kde sa pripája list na
stonku, kým články sú bezlisté úseky medzi dvoma nad sebou stojacimi
uzlami (Bobák et al., 1992).
Obr.
2.1
Stavba stonky (upravené podľa deReffye et al., 1988).
Nové dcérske stonky (konáre) vyrastajú
z púčikov materskej stonky. Púčiky
sú rastové vrcholy s hrbolčekovitými základmi ďalších stoniek
a listov a proti nepriaznivým poveternostným vplyvom ich obaľujú
krycie šupiny. Len výnimočne sú púčiky drevín nahé (kalina
siripútková) alebo polonahé (baza čierna). Púčiky sú na koncoch
stoniek (vrcholové, terminálne) alebo vyrastajú na bokoch, prípadne
v pazuchách listov (bočné, pazušné). Krytosemenné rastliny majú
pazušný púčik takmer za každým listom, nahosemenné (ihličnaté) len
pri malom počte listov. Púčiky majú nielen ochrannú funkciu a sú
základmi nových stoniek, ale už z ich postavenia na stonke možno
usudzovať na stratégiu obsadzovania priestoru, teda rozkonárovania. Len
vďaka rozkonárovaniu môžu stromy ako dlhoveké rastliny najlepšie
využiť priestor a najvhodnejšie nastaviť listy slnečným
lúčom. Púčiky sú tiež dôležitým znakom pri určovaní drevín
v zime, keď nemajú listy, pretože sú pre väčšinu rodov
a druhov charakteristické. Na stonke sú postavené v podstate tromi
spôsobmi: striedavo, protistojne a praslenovito.
Okrem týchto púčikov sa takmer všade na
rastlinnom tele môžu tvoriť vedľajšie (adventívne) púčiky. Pri
drevinách sú to spiace púčiky, ktoré sa prebúdzajú napríklad pri poranení
stromu, alebo vytvárajú charakteristické výmladky.
Nové výhonky
(konáre s listami a pazušnými púčikmi) však nevyrastajú zo
všetkých založených púčikov. Tie, ktoré nevypučali sa môžu
zmeniť na spiace púčiky.
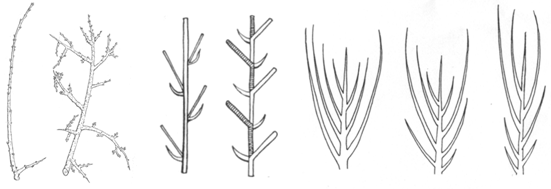
Pri väčšine drevín sa vyskytujú dva základné
typy konárov (Obr. 2.2), ktorých vzájomný pomer a postavenie má
prvoradý vplyv na architektúru koruny.
·
Predlžujúce sa bočné konáriky
(makroblasty) sú dlhé výhonky
s dlhotrvajúcim rastom, dlhými internódiami (článkami) a dlhými
odstupmi medzi bočnými púčikmi. Tvoria sa hlavne z vrcholových
a len ojedinele z bočných púčikov. Takmer všetky mladé
dreviny a niektoré dreviny i v dospelosti (prútnatec, niektoré
vŕby) vytvárajú len tento typ konárov.
·
Skrátené bočné konáriky,
konáriky obmedzeného rastu (brachyblasty)
majú veľmi krátke internódiá, dlhé často len niekoľko desatín
milimetra. Ich povrch je preto nápadne hrboľatý (ginko), čo spôsobujú
jazvy po opadaných krycích šupinách a listové stopy. Najčastejšie
majú len vrcholový púčik, z ktorého vyrastá opäť krátke
internódium. Brachyblasty sú viacročné konáriky s kratšou dobou
trvania (desať až pätnásť rokov) ako pri makroblaste. Sú často
špecializované na určitú funkciu: nesú zväzočky ihlíc (smrekovec,
céder, borovica), kvety (jabloň, hruška) alebo sú premenené na tŕne
(trnka, gledíčia).
Sústava výhonkov sa všeobecne rozkonáruje podľa dvoch typov:
·
Monopodiálne rozkonárovanie – vrcholový
púčik zabezpečuje pokračovanie materskej stonky, z ktorej
vyrastá zreteľná hlavná stonka. Na nej z bočných púčikov
vyrastajú slabšie bočné stonky, ktoré nerastú v smere pôvodnej stonky
a hlavnú stonku ani neprerastajú (ihličnaté dreviny).
·
Sympodiálne rozkonárovanie – rast hlavnej
stonky je prerušený a v jej smere vyrastá dcérska stonka, ktorá sa
vytvorila z najbližšie postaveného pazušného púčika alebo dcérska
stonka natoľko zosilnie, že zatlačí materskú stonku nabok a sama
rastie v smere materskej stonky (dcérske stonky sú hrubšie a dlhšie
ako materská stonka). Výsledné rozkonárenie vypadá navonok ako monopódium, ale
hlavnú stonku tvoria v skutočnosti postupne za sebou nasledujúce
stonky vyššieho stupňa (lipa, brest alebo hrab).
Na prvý pohľad sa oba typy odlíšia podľa
postavenia listov a bočných konárov: zatiaľ čo pri
monopódiu vyrastajú bočné stonky v pazuchách listov, pri sympódiu
vyrastajú „bočné“ (v skutočnosti zatlačené hlavné) stonky proti
listu (Obr.
2.2).
Obr.
2.2
Schematický nákres makroblastu, brachyblastu, monopodiálneho
a sympodiálneho rozkonárovania (Vĕtvička et al., 1992) a
pozdĺžnej symetrie: akrotónia, bazitónia, mezotónia (zľava doprava)
(Bobák et al., 1992).
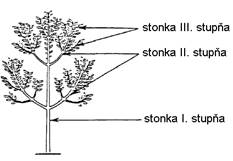
Hlavná stonka sa označuje ako stonka I.
stupňa, na ktorej pri rozkonárovaní vznikajú konáre, stonky II.
stupňa, rozkonárovaním ktorých sa vytvárajú stonky III. stupňa (Obr. 2.3) (Bobák et al., 1992).
Obr.
2.3
Stupeň stonky (upravené podľa deReffye et al., 1988).
Mohutnejším rozvojom konárov v rôznych
výškach materského výhonku sa vytvára pozdĺžna (longitudinálna) symetria
rastlín (Obr.
2.2), a to v prípade monopodiálneho aj
sympodiálneho rozkonárovania. Tým rastliny ako celok nadobúdajú rozmanitý
vzhľad (habitus). Rozlišujeme tri typy symetrie (Bobák et al., 1992):
·
Akrotónia – bočné konáre sa v hornej
časti výhonku vyvíjajú mohutnejšie (stromy).
·
Bazitónia – silnejšie sa vyvíjajú konáre pri
báze stonky (kry). Často sa stáva, že konáre, ktoré sa vyvinuli
z bazálnych púčikov sú oveľa mohutnejšie ako hlavný výhonok a jeho
vyššie konáre, alebo je hlavný výhonok vo svojom vývoji úplne potlačený.
·
Mezotónia – silnejšie sa vyvíjajú stredné
konáre. Tento spôsob rozkonárovania je veľmi zriedkavý.
Rozkonárenie stoniek a vzájomný pomer
konárikov rozličných typov, uhol konárov a vzájomný pomer hrúbky
konárov vyššieho a nižšieho stupňa určujú konečný
vzhľad a textúru dreviny, prípadne jej koruny. Z hľadiska
dreviny je to vždy ukážka snahy organizmu zachytiť čo najviac svetla
i v mnohoposchodovej korune.
Okrem anatomických a morfologických
vlastností ovplyvňujú tvar koruny do istej miery aj vonkajšie
činitele, predovšetkým svetlo a vietor. Suchozemské rastliny
a dreviny osobitne sú prevažne slnné. Rôzne svetelné podmienky (napríklad
zatienenie alebo naopak prebytok svetla) vedú k deformácii „ideálnych“
korún: napríklad jednostranne vyklonené koruny listnatých drevín na okraji lesa
alebo nad hladinou vodnej plochy, kde rastlina reaguje okrem iného aj na
množstvo svetla odrazeného od hladiny. Koruna mnohých drevín vytvára zaoblenú klenutú
plochu (nazýva sa solárna čiže heliodynamická), ktorá vzniká reakciou
rastúcej rastliny na dopad svetelných lúčov a vytvára optimálne
podmienky na využitie svetla. Ak dreviny rastú v malej skupine, reagujú na
slnečné žiarenie spoločne a vytvárajú tiež spoločnú solárnu
plochu, akoby išlo o jednu drevinu. Pretože efekt dopadajúceho
slnečného žiarenia sa mení so zemepisnou šírkou (podľa uhla dopadu
slnečných lúčov), mení sa aj klenba a uhol naklonenia solárnej
plochy (s rastúcou zemepisnou šírkou rastie aj uhol naklonenia solárnej
plochy smerom na slnečnú stranu, klenba je asymetrická, časť na
slnečnej strane je vyvinutejšia ako časť na odvrátenej strane).
K deformácii ideálne stavanej koruny
prispieva okrem svetla aj vietor (najmä vietor vanúci jedným smerom, ako
napríklad na morských brehoch alebo vo vysokých horách, kde vznikajú
zástavovité koruny), zatienenie stavbami alebo stromami, mechanický vplyv
(konkurencia) okolitých drevín. Na stavbu a tvar koruny môže mať
vplyv aj uhol svahu, na ktorom drevina rastie, štruktúra pôdy, prípadne odraz
slnečných slnečných lúčov od nej.
Väčšina zmienených deformácií sa týka len
osamelo rastúcich drevín. Keď drevina rastie v trvalom zápoji,
reaguje príslušnou zmenou tvaru koruny najmä na konkurenciu okolostojacich drevín,
pričom sa spomenuté deformačné vplyvy i vrodené vlastnosti
stavby koruny stávajú druhoradými.
Korunu
stromu tvorí sústava konárov, ktoré vyrastajú z kmeňa:
·
Štrukturálne čiže kostrové
konáre, rozhodujúce pre tvar (skelet) koruny. Sú to väčšie, hrubšie
konáre, obyčajne predlžujúce sa výhonky s vegetatívnou funkciou.
·
Vyplňujúce konáre, a to
slabšie predlžujúce sa výhonky alebo skrátené brachyblasty, ktoré
vypĺňajú korunu a majú rozhodujúce postavenie na obvode (plášti)
koruny; sú to výhonky s vegetatívnou i generatívnou funkciou.
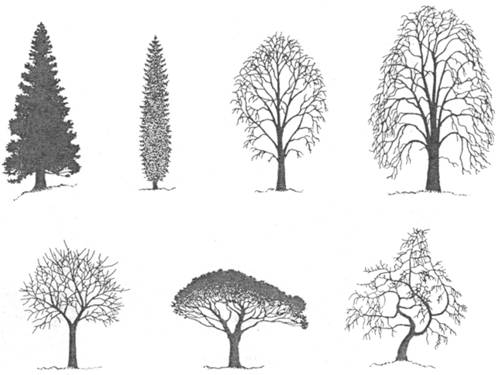
Pre jednotlivé druhy je charakteristická hustota
aj tvar koruny. Ten môžeme pripodobniť ku geometrickým telesám
(kužeľovitá, valcovitá, vretenovitá, valcovitá, elipsoidná, guľovitá)
alebo vyjadriť pomocou špeciálnych termínov (dáždnikovitá, smútočná,
zástavovitá, poschodovitá, nepravidelná) (Obr.
2.4).
Obr.
2.4
Tvar koruny: kužeľovitá, úzko vretenovitá, elipsoidná, vajcovitá,
guľovitá, dáždnikovitá, nepravidelná (zľava hore) (Vĕtvička
et al., 1992).
Dreviny mierneho pásma nemajú veľmi
komplikovanú stavbu koruny, aj keď sa jednotlivé druhy na prvý pohľad
odlišujú. Najväčší rozvoj dosiahla stavba koruny pri drevinách
v tropických zemepisných šírkach. Odtiaľ čerpali Hallé, Oldeman
a Tomlinson inšpiráciu pre svoj klasifikačný
systém výstavby stromových korún (Hallé et al., 1978).
Východiskom boli jednoduché kritériá, ako
napríklad, či výhonky sú rozkonárené (polyaxiálne
dreviny) alebo nerozkonárené (monoaxiálne
dreviny), či ide o monopódium alebo sympódium, či drevina
rastie periodicky, rytmicky (rast prerušovaný obdobím odpočinku) alebo
nepretržito (kontinuálne), či sa korene ďalej rozkonárujú, aké je
postavenie súkvetí a kvetov (terminálne alebo laterálne) atď. Treba
si uvedomiť, že každá z týchto vlastností vstupuje do kombinácie
s ďalšími. Tak vznikla škála dvadsiatich štyroch modelov drevín (Obr. 2.5), nazývaných podľa osôb (napríklad Cookov model,
Prévostov model). Aj keď ide pri tomto spracovaní architektúry drevín
prevažne o tropické dreviny, niektorým modelom zodpovedajú aj dreviny
mierneho pásma; napríklad rad ihličnatých drevín (Araucaria exelsa, jedle, tisy) vyhovuje Massartovmu modelu, Araucaria araucana a väčšina
borovíc vrátane borovice lesnej Rauhovmu modelu, rovnako ako niektoré listnaté
dreviny (javory, duby, jasene), z ďalších listnatých drevín,
napríklad buky, lipy a bresty vyhovujú Trollovmu modelu.
Obr.
2.5
Architektonické modely drevín (Environmental Information Laboratory, 2007).
Dôležitým orgánom drevín sú tiež korene, ktoré sa takisto riadia
určitými zákonitosťami, a vonkajšie prostredie ich deformuje
podstatne výraznejšie ako napr. korunu stromu.
Trojrozmerný
počítačový model nejakého objektu môžeme získať tromi spôsobmi
(Žára et al., 2004):
1)
Obrazovo-orientované
modelovanie (image-based modeling) – ide o získanie trojrozmernej
snímky objektu, teda hrubého modelu získaného priamo z geometrie reálneho
objektu. Tú môžeme vytvoriť pomocou priestorového skeneru alebo
rekonštrukciou z niekoľkých snímkov z fotoaparátu. Nevýhody sú
dané veľkosťou snímaného objektu, prípadne jeho zložitosťou.
2)
Interaktívne
modelovanie (interactive modeling) – model vytvorí človek
(animátor) za pomoci myši, klávesnice, či ďalšieho vstupného
zariadenia. Tento postup je veľmi prácny, vyžaduje talent
a skúsenosti, na druhej strane, výsledok je sémanticky úplne presný.
Animátor totiž vie, čo robí, čo má v úmysle modelovať
a čo ktorá časť modelu znamená. To je v prípade
skenovania neľahké.
3)
Procedurálne
modelovanie (procedural modeling) – pôvodné geometrické modely
(polygonálne modely, krivky, body, čiary) sú nepostačujúce na to, aby
reprezentovali komplexné objekty. Preto boli vyvinuté modelovacie techniky,
ktoré umožnili abstrakciu modelu, lepšiu špecifikáciu alebo vyššiu kontrolu nad
modelom. Väčšina týchto techník je vo svojej podstate procedurálna.
Procedurálne techniky sú časti kódu alebo algoritmy, ktoré presne
určujú niektoré charakterisktiky počítačového modelu alebo
efektu (Ebert et al., 1998). Model je teda získaný generovaním pomocou nejakého
algoritmu. Túto metódu možno rozdeliť na dve základné podtriedy. Prvou
z nich sú metódy, používané v CAD a CAGD ako napríklad šablónovanie a
generovanie plôch z kriviek. Druhá podtrieda sa zameriava na automatické
generovanie objektov, ktoré sa vizuálne alebo správaním podobajú objektom,
s ktorými sa stretávame v prírode. Túto podtriedu môžeme
rozdeliť na ďalšie skupiny:
4) Algoritmy, ktoré vychádzajú
z gramatík – napríklad L-systémy, používané najčastejšie na
generovanie rastlín. Pomocou niekoľkých parametrov umožňujú
simulovať zložité modely stromov, rastlín alebo iných prírodných objektov.
5) Fraktálna geometria – algoritmy na
generovanie hôr, krajín, kameňov, koralov, stromov atď.
6) Systémy častíc – systém
častíc sa reprezentuje ako veľký súbor veľmi malých prvkov
(častíc), ktorých vlastnosti sa stochasticky menia v čase. Preto
systémy častíc používajú veľkú databázu geometrických prvkov, ktoré
reprezentujú prírodné objekty, ale animácia, pozícia, narodenie a smrť
častíc sa kontrolujú algoritmicky. Patria tu algoritmy na generovanie
napr. explózií, kŕdľa vtákov, padajúcich lôpt, ekosystémov, simuláciu
ohňa, dymu a pod.
Výhodou
procedurálnych metód je tzv. database amplification factor (Smith,
1984), ktorý umožňuje generovať a kontrolovať širokú škálu
komplexných štruktúr (databázu častíc) na základe malého počtu
parametrov.
Kým umelo
vytvorené predmety sa vyznačujú presnosťou geometrického opisu, popis
prírodných tvarov technikami klasickej geometrie je neefektívny. Ťažko
popísať mrak ako množinu gulí alebo parametrickú plochu, či
modelovať krajinu ako množinu trojuholníkov. Silný a relatívne
jednoduchý aparát, ktorý takéto modelovanie umožňuje, poskytuje fraktálna
geometria. Na rozdiel od euklidovskej geometrie, v ktorej popisujeme
objekty pomocou rovníc, vo fraktálnej geometrii sa používajú na popis objektov
algoritmy, najčastejšie rekurzívne. Kým klasická geometria popisuje
invariancie vzhľadom k transformáciám, ako sú otočenie či
posunutie, fraktálna geometria sa zaoberá invarianciou voči zmene mierky
(Žára et al., 2004). Za otca tejto vednej disciplíny sa považuje Benoit B.
Mandelbrot (Peitgen et al., 1992) (Obr.
4.1).
Obr. 4.1 Mandelbrotova množina (Mandelbrot set,
2007).
Hlavným pojmom fraktálnej
geometrie je samopodobnosť (self-similarity).
Samopodobnú štruktúru môžeme rozdeliť na štruktúry, z ktorých každá
je zmenšenou kópiou originálu.
Mandelbrot
definoval fraktál (z lat. frangere,
zlomiť) ako množinu, ktorej Hausdorff-Besicovitchova dimenzia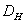 je ostro väčšia ako topologická dimenzia
je ostro väčšia ako topologická dimenzia 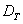 (Mandelbrot, 1982).
Matematicky je fraktál popísaný ako množina, ktorá vznikne zlúčením
nekonečne veľa kópií samej seba (4.1) (Žára et al., 2004, str. 269):
(Mandelbrot, 1982).
Matematicky je fraktál popísaný ako množina, ktorá vznikne zlúčením
nekonečne veľa kópií samej seba (4.1) (Žára et al., 2004, str. 269):
 . (4.1)
. (4.1)
Objekty, ktoré
pozorujeme v prírode, však s veľkou pravdepodobnosťou
nemajú nekonečné hierarchie detailov. V zmysle rovnice sa teda
nejedná o fraktály, napriek tomu sa však na základe všeobecnej dohody za
fraktály považujú, aj keď presnejšie by bolo označenie fraktálom
podobné. Veľkú väčšinu členitých objektov, s ktorými sa
stretávame v prírode, môžeme popísať pomocou fraktálov.
Aj u rastlín
môžeme pozorovať samopodobnosť, čo nám umožňuje abstrakciu
opakujúcich sa základných rastlinných štruktúr (modulov)
a uľahčuje prácu pri pochopení stavby rastlín, ich analýze
a simulácii. Niekedy sa moduly spájajú do zložitejších útvarov,
v ktorých sa podobné štruktúry objavujú na rôznych úrovniach. Napríklad
kvety usporiadané v zložených súkvetiach alebo viacnásobne zložené listy
rastlín, ktorých lístky sú podobné celému listu (Obr. 4.2). Tieto vlastnosti využíva pri modelovaní
aj Prusinkiewicz (2001).
Obr. 4.2 Ukážky samopodobnosti u rastlín.
Samopodobnosť
možno pozorovať aj u stromov, napr. pri rozkonárovaní, aj keď je
menej výrazná ako u bylinných druhov. Je to spôsobené tým, že ich rast trvá
veľmi dlho (v porovnaní s bylinnými druhmi), a preto sú
viac ovplyvnené vonkajšími vplyvmi prostredia.
Napriek tomu, že
popis rastlín ako celku, zloženého z podobných častí, má veľký
význam (pre ich analýzu a následné modelovanie) a vnímanie samopodobnosti u
rastlín je známe už niekoľko desaťročí, doteraz neexistovalo
žiadne formálne kritérium pre určenie stupňa podobnosti.
Prusinkiewicz (2004b) predstavil nový pojem, topologickú samopodobnosť (topological
self-similarity), ktorá je u rastlín bežnejšia ako geometrická samopodobnosť (geometric
self-similarity). Geometrická samopodobnosť sa vo všeobecnosti
vyjadruje ako množina afinných transformácií, ktoré zobrazujú štruktúru do jej
zložiek, kým topologická sa zaoberá usporiadaním a vzťahmi medzi
týmito zložkami. Formálnu definíciu samopodobnosti a jej merania,
prispôsobenú na vetviace sa rastlinné štruktúry, navrhli Ferraro et al. (2005).
Na jej meranie používajú algoritmus na výpočet topologických vzdialeností
medzi rozkonárenými štruktúrami.
Aký je však
vzťah medzi fraktálmi a rastlinami? Považovať rastliny za
aproximácie „perfektných“ fraktálnych štruktúr by nebolo akceptovateľné.
Životaschopnejší prístup je ten opačný, a to považovať fraktály za
abstraktný popis reálnych biologických štruktúr (Prusinkiewicz et al., 1990).
Môže sa zdať, že popísať list pomocou hladkej krivky je jednoduchšie
ako pomocou nepravidelného obrysu fraktálu. Vysvetlenie spočíva v tom,
čo znamená jednoduchšie. Intuitívne sa krivka zdá byť jednoduchšia,
ale ak meriame zložitosť objektu dĺžkou najkratšieho algoritmu, ktorý
ho generuje, opak je pravdou.
Fraktálny prístup
k analýze rastliny, s dôrazom na samopodobnosť, ponúka
kľúč k pochopeniu komplexnej stavby rastlín a ponúka
rekurzívne procedúry, cez ktoré mohli byť jednotlivé štruktúry vytvorené.
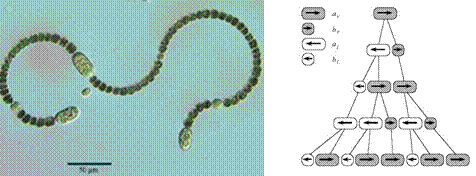
V roku 1968 biológ Aristid Lindenmayer
predstavil formalizmus na modelovanie a simulovanie vývoja mnohobunkových
organizmov (sinice Anabaena catenula)
(Obr. 5.1), neskôr nazvaný L-systémy (Lindenmayerove systémy).
L-systémy pôvodne neumožňovali modelovanie
vyšších rastlín. Neskôr však bolo navrhnutých niekoľko geometrických
interpretácií L-systémov, vďaka ktorým sa z nich stal univerzálny
nástroj na modelovanie rastlín a boli prijaté ako matematický formalizmus pre
popis vývinu rastlín (Prusinkiewicz, 1990, Prusinkiewicz et al., 1995).
L-systémy možno chápať ako
modifikáciu formálnych jazykov, keďže ich základným konceptom je
prepisovanie reťazcov (postupnosti symbolov) (string-rewriting systems) podľa určitých pravidiel. Pri
tomto postupe sa zložité objekty definujú pomocou postupne sa nahrádzajúcich
častí počiatočného objektu. Získaná postupnosť sa potom
interpretuje geometricky. Rozdiel medzi formálnymi jazykmi a L-systémami
je v spôsobe aplikácie prepisovacích pravidiel. Pri formálnych gramatikách
sú pravidlá aplikované postupne, kým pri L-systémoch sú použité súbežne
(paralelne). Tento rozdiel vyplýva z biologickej motivácie L-systémov
(pravidlá majú zachytiť delenie buniek v mnohobunkovom organizme, kde
sa súčasne delí viacero buniek naraz).
Najjednoduchšiu triedu L-systémov predstavujú
deterministické bezkontextové L-systémy,
označované ako D0L-systémy.
Formálne ich môžeme definovať takto (Prusinkiewicz, 1990):
Nech 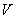 je abeceda,
je abeceda, 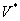 je množina všetkých
slov nad a
je množina všetkých
slov nad a 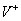 je množina neprázdnych
slov nad V. Reťazcový 0L-systém je usporiadaná trojica
je množina neprázdnych
slov nad V. Reťazcový 0L-systém je usporiadaná trojica 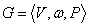 , kde:
, kde:
·
je abeceda systému,
·
 je neprázdne slovo
nazývané axióma a
je neprázdne slovo
nazývané axióma a
·
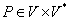 je konečná množina prepisovacích pravidiel.
je konečná množina prepisovacích pravidiel.
Prepisovacie pravidlo  budeme zapisovať
ako
budeme zapisovať
ako 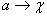 . Písmeno
. Písmeno 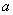 sa nazýva predchodca (predecessor, ľavá strana, prepisovaný symbol) a slovo
sa nazýva predchodca (predecessor, ľavá strana, prepisovaný symbol) a slovo  sa nazýva nasledovník (successor, pravá strana) pravidla. Predpokladá sa, že pre každé
písmeno
sa nazýva nasledovník (successor, pravá strana) pravidla. Predpokladá sa, že pre každé
písmeno 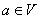 existuje aspoň
jedno slovo
existuje aspoň
jedno slovo 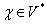 také, že . Ak nie je pre niektorý prepisovaný symbol explicitne zadané
prepisovacie pravidlo, predpokladá sa, že
také, že . Ak nie je pre niektorý prepisovaný symbol explicitne zadané
prepisovacie pravidlo, predpokladá sa, že 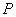 obsahuje identické pravidlo
obsahuje identické pravidlo 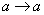 . 0L-systém je deterministický (označenie D0L-systém), práve vtedy, keď pre
každé existuje práve jedno také, že .
. 0L-systém je deterministický (označenie D0L-systém), práve vtedy, keď pre
každé existuje práve jedno také, že .
Nech  je ľubovoľné
slovo nad abecedou V. Slovo
je ľubovoľné
slovo nad abecedou V. Slovo  je priamo odvodené zo slova
je priamo odvodené zo slova 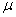 (alebo generované slovom ), označenie
(alebo generované slovom ), označenie 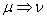 , práve vtedy, keď
, práve vtedy, keď 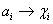 , pre všetky
, pre všetky 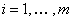 . Slovo
. Slovo 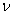 je generované
L-systémom
je generované
L-systémom 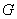 deriváciou dĺžky n, ak existuje derivačná postupnosť slov
deriváciou dĺžky n, ak existuje derivačná postupnosť slov  taká, že
taká, že 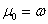 ,
, 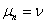 a
a 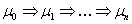 .
.
Nasledujúci príklad ilustruje použitie
D0L-systémov na simuláciu rastu časti mnohobunkového vlákna sinice Anabaena catenula. Symboly a  reprezentujú
cytologické štádium buniek (ich veľkosť a schopnosť
deliť sa). Indexy
reprezentujú
cytologické štádium buniek (ich veľkosť a schopnosť
deliť sa). Indexy 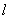 a
a  udávajú polaritu
bunky, ktorá určuje pozíciu, v ktorej budú vytvorené dcérske bunky
typu alebo . Vývoj je popísaný nasledujúcim L-systémom:
udávajú polaritu
bunky, ktorá určuje pozíciu, v ktorej budú vytvorené dcérske bunky
typu alebo . Vývoj je popísaný nasledujúcim L-systémom:
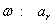
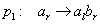
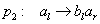

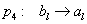
Začínajúc jednou bunkou 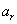 (axióma)
a použitím prepisovacích pravidiel
(axióma)
a použitím prepisovacích pravidiel 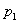 až
až 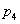 , vzniká nasledujúca postupnosť slov:
, vzniká nasledujúca postupnosť slov:
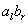
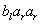

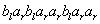

Pod mikroskopom by sme mohli filamenty (vlákna)
pozorovať ako sled valcových útvarov (buniek) rôznej dĺžky,
s dlhšími bunkami typu a kratšími
bunkami typu (Obr. 5.1). Vzhľadom na diskrétny charakter D0L-systémov,
rast buniek medzi deleniami nie je zachytený.
Obr.
5.1
Vlákno sinice a simulácia vývinu vlákna použitím D0L-systému
(Prusinkiewicz et al., 1990).
V snahe použiť L-systémy na modelovanie
vyšších rastlín bolo navrhnutých niekoľko grafických interpretácií
L-systémov. Ich prehľad možno nájsť napr. v (Prusinkiewicz et al.,
1990, s. 6). Prusinkiewicz sa zameral na interpretáciu pomocou korytnačej
grafiky, známej z jazyka LOGO (Abelson et al., 1982). Základná myšlienka spočíva v tom, že reťazec
symbolov vytvorený pomocou L-systému sa interpretuje ako postupnosť
príkazov, ktoré kontrolujú pohyb korytnačky (Prusinkiewicz, 1986a).
Stav korytnačky (state of the turtle) je definovaný ako usporiadaná trojica 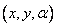 , kde karteziánske súradnice
, kde karteziánske súradnice  reprezentujú pozíciu korytnačky a uhol
reprezentujú pozíciu korytnačky a uhol  jej orientáciu (smer, v ktorom sa
korytnačka pohybuje dopredu). Na základe vstupných údajov, ktorými sú dĺžka kroku d a prírastok uhla
jej orientáciu (smer, v ktorom sa
korytnačka pohybuje dopredu). Na základe vstupných údajov, ktorými sú dĺžka kroku d a prírastok uhla 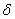 , môže korytnačka odpovedať na príkazy, ktoré sú
reprezentované nasledujúcimi symbolmi (Obr.
5.2):
, môže korytnačka odpovedať na príkazy, ktoré sú
reprezentované nasledujúcimi symbolmi (Obr.
5.2):
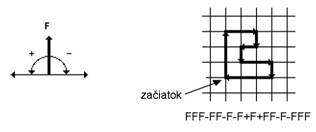
F Pohni
sa dopredu o krok dĺžky d
a nakresli čiaru zo starej do novej pozície. Stav korytnačky sa
zmení na 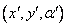 , kde
, kde 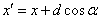 a
a 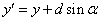 .
.
f Pohni
sa dopredu o krok dĺžky d,
bez nakreslenia čiary.
+ Otoč
sa doľava o uhol (proti smeru
hodinových ručičiek). Stav korytnačky sa zmení na 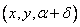 .
.
 Otoč sa doprava
o uhol (v smere hodinových
ručičiek). Stav korytnačky sa zmení na
Otoč sa doprava
o uhol (v smere hodinových
ručičiek). Stav korytnačky sa zmení na  .
.
Stav korytnačky sa zmení zakaždým, keď
sa interpretuje symbol. Ak je daný reťazec , pôvodný stav korytnačky 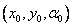 a pevné parametre d a , odpoveď korytnačky na reťazec sa interpretuje ako
postupnosť korytnačkou nakreslených čiar (Obr. 5.2).
a pevné parametre d a , odpoveď korytnačky na reťazec sa interpretuje ako
postupnosť korytnačkou nakreslených čiar (Obr. 5.2).
Obr.
5.2
Interpretácia symbolov 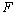 ,
,  ,
, 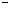 pomocou
korytnačej grafiky (Prusinkiewicz et al., 1990).
pomocou
korytnačej grafiky (Prusinkiewicz et al., 1990).
L-systémy možno použiť aj na generovanie
klasických fraktálov (Prusinkiewicz et al., 1990, Peitgen et al., 1992).
Napríklad všeobecne známu Kochovu vločku (Obr. 5.3) možno získať interpretáciou reťazcov
vytvorených nasledujúcim L-systémom:
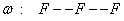


Obr.
5.3
Prepisovacie pravidlo p (hore), axióma ω (dole vľavo)
a niekoľko prvých iterácií Kochovej vločky (Prusinkiewicz et
al., 2003).
Interpretáciu L-systémov pomocou korytnačej
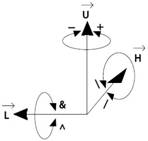
grafiky možno rozšíriť do priestoru (Prusinkiewicz et al., 1990). Aktuálna
orientácia korytnačky v priestore je daná tromi vektormi 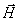 ,
,  ,
,  , ktoré označujú:
, ktoré označujú:
(heading) smer, v ktorom sa korytnačka pozerá dopredu,
(left) smer doľava a
(up) smer hore.
Obr.
5.4
Kontrola korytnačky v priestore (Prusinkiewicz et al., 1990).
Tieto vektory majú jednotkovú dĺžku, sú
navzájom kolmé a spĺňajú rovnosť 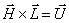 . Rotácie korytnačky možno potom vyjadriť
rovnosťou
. Rotácie korytnačky možno potom vyjadriť
rovnosťou 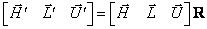 , kde
, kde  je rotačná matica
rozmeru
je rotačná matica
rozmeru 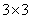 . Rotácie o uhol okolo vektorov , , sa reprezentujú
maticami:
. Rotácie o uhol okolo vektorov , , sa reprezentujú
maticami:
 (5.1)
(5.1)
 (5.2)
(5.2)
 (5.3)
(5.3)
Pôvodný stav korytnačky je daný v
počiatku kateziánskej súradnicovej sústavy, s vektorom v kladnom smere osi y a vektorom v zápornom smere osi x.
Orientáciu korytnačky v priestore
riadime nasledujúcimi symbolmi (Obr.
5.4):
 Otoč sa
doľava (turn left) o uhol . Použije sa rotačná matica
Otoč sa
doľava (turn left) o uhol . Použije sa rotačná matica  .
.
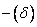 Otoč sa
doprava (turn right) o uhol . Použije sa rotačná matica
Otoč sa
doprava (turn right) o uhol . Použije sa rotačná matica 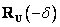 .
.
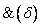 Nakloň sa
dolu (pitch down) dolu o uhol . Použije sa rotačná matica
Nakloň sa
dolu (pitch down) dolu o uhol . Použije sa rotačná matica 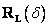 .
.
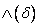 Nakloň sa
hore (pitch up) o uhol . Použije sa rotačná matica
Nakloň sa
hore (pitch up) o uhol . Použije sa rotačná matica 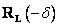 .
.
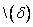 Nakloň
sa doľava (roll left)
o uhol . Použije sa rotačná matica
Nakloň
sa doľava (roll left)
o uhol . Použije sa rotačná matica 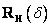 .
.
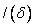 Nakloň
sa doprava (roll right) o uhol . Použije sa rotačná matica
Nakloň
sa doprava (roll right) o uhol . Použije sa rotačná matica  .
.
 Otoč sa
dookola. Použije sa rotačná matica
Otoč sa
dookola. Použije sa rotačná matica  .
.
Podrobnejšie spracovanú grafickú interpretáciu
L-systémov pomocou korytnačej grafiky s mnohými ďalšími
rozšíreniami, výrazne zlepšujúcimi realizmus modelov možno nájsť v (Mĕch
et al., 2007).
Podľa doteraz predstavených pravidiel
korytnačka interpretuje reťazec symbolov ako postupnosť
čiar. V závislosti od ich dĺžky a uhlov medzi nimi sa
výsledná čiara pretína alebo nie, je viac či menej zatočená
a môže mať niektoré segmenty nakreslené viackrát alebo iné mať
neviditeľné, stále však ostáva iba jednoduchou čiarou. V prírode
ale dominujú vetviace sa štruktúry, a preto bol, na modelovacie účely,
potrebný aj matematický popis tvarov majúcich „stromovitú“ štruktúru
a metódy na ich generovanie. Jedna z možností je reprezentovať
rozkonárené časti ako reťazce so zátvorkami [ a ], ktoré budú
vymedzovať určitý konár stromu, teda budú predstavovať jeho
začiatok a koniec (Obr. 5.5).
Obr.
5.5
Príklady rozvetvených štruktúr vytvorených zátvorkovými L-systémami
(Prusinkiewicz et al., 1990).
Korytnačke do funkčnosti pribudne
zásobník, do ktorého bude môcť ukladať svoj stav (Prusinkiewicz et
al., 1990). V korytnačej grafike sa dva nové symboly interpretujú
takto:
[ Ulož
do zásobníka aktuálny stav korytnačky. Do zásobníka sa uloží informácia o
pozícii a orientácii korytnačky a tiež tu môžu byť údaje
o iných atribútoch, napríklad o farbe a šírke kreslených
čiar.
] Vyber
stav korytnačky zo zásobníka a nastav ho ako aktuálny. Nenakreslí sa
žiadna čiara aj keď sa pozícia korytnačky obyčajne mení.
Hlavným problémom pri D0L-systémoch je, že
dĺžka čiar sa redukuje na celočíselné násobky jednotkového
segmentu. Okrem toho každá zmena dĺžky čiary alebo uhla otočenia
by znamenala nový symbol, čo by viedlo k veľkému nárastu symbolov a
prepisovacích pravidiel a špecifikácia D0L-systému by sa príliš skomplikovala.
Tento problém bol vyriešený zavedením parametrických
L-systémov (Prusinkiewicz et al., 1997), ktoré umožňujú doplniť
jednotlivé symboly o číselné parametre (Obr.
5.6).
Parametrické L-systémy pracujú s parametrickými slovami, čo sú
reťazce modulov, skladajúcich sa
z písmen abecedy a k
nim priradených parametrov.
Písmená sú prvky abecedy a parametre sú
z množiny reálnych čísel 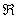 . Modul s písmenom
. Modul s písmenom  a parametrami
a parametrami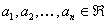 je označený ako
je označený ako  . Každý modul je z množiny
. Každý modul je z množiny 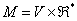 , kde
, kde 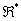 je množina všetkých
konečných postupností parametrov. Množina všetkých reťazcov modulov
a množina všetkých neprázdnych reťazcov je označená ako
je množina všetkých
konečných postupností parametrov. Množina všetkých reťazcov modulov
a množina všetkých neprázdnych reťazcov je označená ako 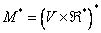 a
a  .
.
Modul má vo svojej špecifikácii formálne parametre, ktoré nadobúdajú konkrétne
hodnoty (aktuálne parametre)
z množiny reálnych čísel. Aktuálne
parametre, ktoré sa nachádzajú v slovách, sa zhodujú s formálnymi parametrami, ktoré sú použité
v prepisovacích pravidlách L-systémov. Ak  je množina formálnych parametrov, potom
je množina formálnych parametrov, potom  označuje logický výraz s parametrami z a
označuje logický výraz s parametrami z a  je aritmetický výraz s parametrami
z tej istej množiny. Obidva typy výrazov sa skladajú z formálnych
parametrov a číselných konštánt, ktoré sú kombinované použitím
aritmetických operátorov
je aritmetický výraz s parametrami
z tej istej množiny. Obidva typy výrazov sa skladajú z formálnych
parametrov a číselných konštánt, ktoré sú kombinované použitím
aritmetických operátorov  ,
, 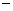 ,
, 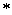 ,
, 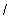 ; exponenciálneho operátora
; exponenciálneho operátora 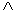 , relačných operátorov
, relačných operátorov  ,
,  ,
,  ,
,  ,
, 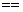 ; logických operátorov
; logických operátorov 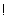 ,
, 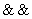 ,
,  (negácia, a, alebo) a
zátvorky (). Výrazy tiež môžu obsahovať volania štandardných matematických
funkcií ako prirodzený logaritmus, sínus, funkcie vracajúce náhodné premenné.
Symboly pre operátory a pravidlá na vytvorenie syntakticky správnych výrazov
sú rovnaké ako pre programovací jazyk C. Relačné a logické výrazy
nadobúdajú pre nepravdivý výrok (false)
hodnotu 0, pre pravdivý výrok (true)
hodnotu 1. Logické tvrdenie označené ako prázdny reťazec má hodnotu
rovnú 1. Množiny všetkých správne vytvorených logických a aritmetických
výrazov s parametrami z sa označujú
(negácia, a, alebo) a
zátvorky (). Výrazy tiež môžu obsahovať volania štandardných matematických
funkcií ako prirodzený logaritmus, sínus, funkcie vracajúce náhodné premenné.
Symboly pre operátory a pravidlá na vytvorenie syntakticky správnych výrazov
sú rovnaké ako pre programovací jazyk C. Relačné a logické výrazy
nadobúdajú pre nepravdivý výrok (false)
hodnotu 0, pre pravdivý výrok (true)
hodnotu 1. Logické tvrdenie označené ako prázdny reťazec má hodnotu
rovnú 1. Množiny všetkých správne vytvorených logických a aritmetických
výrazov s parametrami z sa označujú 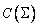 ,
, 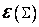 .
.
Parametrický
0L-systém je usporiadaná štvorica 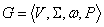 , kde
, kde
·
je abeceda systému,
·
je množina formálnych parametrov,
·
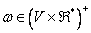 je neprázdne
parametrické slovo nazývané axióma,
je neprázdne
parametrické slovo nazývané axióma,
·
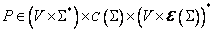 je konečná množina pravidiel.
je konečná množina pravidiel.
Symboly  a
a  sú použité na
oddelenie troch častí pravidla: prepisovaný symbol (predecessor), podmienka (condition),
pravá strana pravidla (successor).
Preto má pravidlo v 0L-systéme tvar
sú použité na
oddelenie troch častí pravidla: prepisovaný symbol (predecessor), podmienka (condition),
pravá strana pravidla (successor).
Preto má pravidlo v 0L-systéme tvar
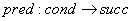 .
.
Číslica „0“ v názve „0L-systém“ znamená,
že nezáleží na kontexte, v akom sa prepisovaný symbol nachádza.
Napríklad pravidlo s prepisovaným symbolom 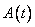 , podmienkou
, podmienkou  a pravou stranou
a pravou stranou 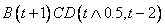 je zapísané ako
je zapísané ako
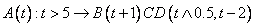 .
.
Prepisovacie pravidlo môže byť aplikované na
modul v parametrickom slove, ak sú splnené nasledujúce podmienky:
·
písmeno v module je rovnaké
ako písmeno v prepisovanom symbole,
·
počet aktuálnych parametrov
v module je rovný počtu formálnych parametrov v predchodcovi
prepisovacieho pravidla a
·
podmienka je pravdivá, ak sú
formálne parametre v pravidle nahradené aktuálnymi parametrami.
Odvodenie je podobné ako pri bezkontextových
L-systémoch.
Pri interpretácii korytnačka číta
jednotlivé moduly a hodnoty ich parametrov interpretuje geometricky.
Parameter môže určovať dĺžku kroku korytnačky, hrúbku
čiary, ktorú nakreslí alebo uhol, o ktorý sa otočí. Ak je
k symbolu priradených viac parametrov, hodnotu stavu korytnačky bude
určovať prvý parameter. Ak sa za symbolom nenachádza žiadny
parameter, budú použité prednastavené hodnoty, špecifikované na začiatku.
Základné symboly, ktoré môžu byť ovplyvnené parametrami, sú tieto:
 Pohni sa
dopredu o krok dĺžky
Pohni sa
dopredu o krok dĺžky  a nakresli
čiaru zo starej do novej pozície. Pozícia korytnačky sa zmení na
a nakresli
čiaru zo starej do novej pozície. Pozícia korytnačky sa zmení na  .
.
 Pohni sa
dopredu o krok dĺžky , bez nakreslenia čiary.
Pohni sa
dopredu o krok dĺžky , bez nakreslenia čiary.
 Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa otočí doľava, ak je záporné,
korytnačka sa otočí doprava.
Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa otočí doľava, ak je záporné,
korytnačka sa otočí doprava.
 Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa natočí dolu, ak je záporné,
korytnačka sa natočí hore.
Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa natočí dolu, ak je záporné,
korytnačka sa natočí hore.
 Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa natočí doprava, ak je záporné,
korytnačka sa natočí doľava.
Otoč sa
okolo o uhol stupňov. Ak je kladné,
korytnačka sa natočí doprava, ak je záporné,
korytnačka sa natočí doľava.
Symboly , 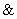 , a sa používajú ako
písmená abecedy a tiež ako
operátory v logických a aritmetických výrazoch, pričom ich
význam závisí od kontextu.
, a sa používajú ako
písmená abecedy a tiež ako
operátory v logických a aritmetických výrazoch, pričom ich
význam závisí od kontextu.
Obr.
5.6
Príklady stromových štruktúr vytvorených parametrickým L-systémom, s rôznymi
hodnotami parametrov pre uhly rozkonárovania, dĺžku a šírku konárov a pod.
(Prusinkiewicz et al., 1997).
Všetky rastliny vytvorené deterministickým
L-systémom sú identické. Premenlivosť, charakteristická pre biologické
organizmy, môže byť dosiahnutá pridaním náhodnosti do interpretácie
symbolov pomocou korytnačky, L-systému alebo oboch (Obr. 5.7). Náhodnosť (randomization)
interpretácie ako samotnej má limitujúci efekt. Kým geometria (geometric aspect) rastliny, ako
napríklad dĺžka stonky alebo uhly medzi vetvami, je zmenená, základná
topológia ostáva nezmenená. Naopak, pridanie náhodnosti do použitia
prepisovacích pravidiel, môže ovplyvniť topológiu aj geometriu rastliny
(Prusinkiewicz et al., 1990).
Parametrický
stochastický 0L-systém je usporiadaná pätica 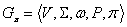 (Prusinkiewicz et al.,
1997). Abeceda V, množina formálnych
parametrov , axióma
(Prusinkiewicz et al.,
1997). Abeceda V, množina formálnych
parametrov , axióma  a množina
prepisovacích pravidiel P sa definujú
ako pri parametrických D0L-systémoch. Funkcia
a množina
prepisovacích pravidiel P sa definujú
ako pri parametrických D0L-systémoch. Funkcia 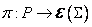 určuje
aritmetický výraz vracajúci nezáporné číslo, nazývané pravdepodobnostný faktor (probability
factor) pre každé prepisovacie pravidlo . Pravidlo v stochastickom L-systéme bude mať tvar
určuje
aritmetický výraz vracajúci nezáporné číslo, nazývané pravdepodobnostný faktor (probability
factor) pre každé prepisovacie pravidlo . Pravidlo v stochastickom L-systéme bude mať tvar
 ,
,
kde pred,
cond a succ majú taký istý význam ako v parametrických L-systémoch
a prob je výraz vracajúci
pravdepodobnostný faktor.
Ak 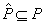 je množina
prepisovacích pravidiel, ktoré možno aplikovať na daný modul
je množina
prepisovacích pravidiel, ktoré možno aplikovať na daný modul 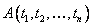 v prepisovanom
reťazci, potom pravdepodobnosť
v prepisovanom
reťazci, potom pravdepodobnosť 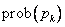 použitia príslušného
pravidla
použitia príslušného
pravidla 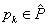 na tento modul je
rovná:
na tento modul je
rovná:
 . (5.4)
. (5.4)
Vo všeobecnosti táto pravdepodobnosť nie je
konštanta priradená k pravidlu, ale závisí od hodnôt parametra
v prepisovanom module.
Odvodenie budeme nazývať stochastická
derivácia v 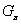 , ak pre každý výskyt modulu v slove je
pravdepodobnosť aplikácie pravidla
, ak pre každý výskyt modulu v slove je
pravdepodobnosť aplikácie pravidla 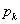 s prepisovaným
symbolom daná rovnicou (5.4)
s prepisovaným
symbolom daná rovnicou (5.4)
Podľa tejto definície, v jednom
derivačnom kroku môžu byť použité rôzne pravidlá s rovnakou
ľavou stranou na rovnaký modul, vyskytujúci sa na rôznych miestach.
Obr.
5.7
Príklady stromových štruktúr vytvorených stochastickým L-systémom (Prusinkiewicz
et al., 1994).
Pravidlá v 0L-systémoch sú bezkontextové, t.
j. sú použité bez ohľadu na to, v akom kontexte sa nachádza
prepisovaný symbol. Aplikácia pravidla závislého od kontextu umožňuje
simulovať interakcie medzi jednotlivými časťami rastliny
a šírenie látok (živín, hormónov) vo vnútri rastliny.
Kontextové
L-systémy môžeme podľa tvaru pravidiel
rozdeliť do niekoľkých tried (Prusinkiewicz et al., 1990).
2L-systémy
používajú pravidlá v tvare 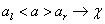 , kde symbol (strict predecessor) môže byť nahradený pravou stranou vtedy
a len vtedy, ak mu predchádza symbol
, kde symbol (strict predecessor) môže byť nahradený pravou stranou vtedy
a len vtedy, ak mu predchádza symbol 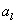 a nasleduje ho
symbol . Symboly a tvoria ľavý a pravý kontext symbolu v tomto pravidle.
Pravidlo v kontextovom L-systéme možno zapísať nasledovne:
a nasleduje ho
symbol . Symboly a tvoria ľavý a pravý kontext symbolu v tomto pravidle.
Pravidlo v kontextovom L-systéme možno zapísať nasledovne:
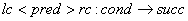 ,
,
kde pred,
cond a succ majú taký istý význam ako v parametrických L-systémoch, lc a rc sú ľavý (left context)
a pravý kontext (right context).
Pravidlá v 1L-systémoch majú len ľavý alebo len pravý kontext, teda sú
tvaru 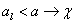 alebo
alebo  .
.
0L-systémy, 1L-systémy a 2L-systémy patria do
širšej triedy IL-systémov, tiež
nazývaných (k,l)-systémy. V
(k,l)-systémoch je ľavý kontext slovo dĺžky 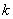 a pravý kontext je slovo dĺžky .
a pravý kontext je slovo dĺžky .
Pôvodný formalizmus L-systémov poskytuje model
vývinu, ktorý je diskrétny v čase aj priestore. Diskretizácia
v čase znamená, že stav modelu je známy iba v presných
časových intervaloch. Diskretizácia v priestore znamená, že rozsah stavov
modelu je limitovaný na konečný počet. Tomuto sa dá vyhnúť
použitím parametrických L-systémov, modely sú však aj napriek tomu stále známe
iba v diskrétnych časových intervaloch. Toto predstavuje problém pri
animáciách, kde sa vyžaduje plynulý sled vývinových foriem. Vývin prebiehajúci
v spojitom čase možno vyjadriť použitím formalizmu časovaných D0L-systémov (Prusinkiewicz
et al., 1990). Základný rozdiel medzi týmto a predchádzajúcimi typmi
L-systémov spočíva v definícii dĺžky derivácie. Kým u
„obyčajných“ L-systémov sa vyjadruje ako počet derivačných
krokov, u časovaných L-systémov predstavuje celkový uplynutý
čas od začiatku pozorovania (derivačného procesu).
Veľa štruktúr v prírode má zložitú
topológiu, ktorú nemožno popísať pomocou doterajších rozšírení
(formalizmu) L-systémov. Medzi ne patria jednovrstvové (mikroskopické) bunkové
štruktúry, napr. papradie gametofyty (štádium životného cyklu, v ktorom sa
tvoria pohlavné bunky, pohlavná generácia), živočíšne embryá, rastlinná
pokožka (Obr. 5.8). Z matematického pohľadu ich
možno reprezentovať pomocou rovinných grafov s cyklami, nazývaných mapy (maps). Mapa je konečná množina regiónov (regions), kde
každý región je obklopený hranicou, ktorá pozostáva z konečnej,
kruhovej postupnosti hrán (edges), stretávajúcich sa vo vrcholoch (vertices). Pre hrany a vrcholy platia určité pravidlá
(Nakamura et al., 1986).
Obr.
5.8
Simulovaný vývoj a fotografia gametofytu druhu Microsorium linguaeforme (Prusinkiewicz et al., 1990).
Mapy sa zhodujú s mikroskopickým
pohľadom na vrstvu buniek. Regióny reprezentujú bunky a hrany bunkové
steny, kolmé na rovinu pohľadu.
Na popis a simuláciu vývinu týchto bunkových
štruktúr slúži nová trieda, mapové L-systémy (Prusinkiewicz et al., 1990) (Obr. 5.9). Mapový
0L-systém je prepisovací systém, ktorý nepracuje nad reťazcami
symbolov, ale nad mapami, pričom neumožňuje interakciu medzi
regiónmi. Región sa môže rozdeliť na najviac dva dcérske regióny, hrany
nemožno zrušiť, čo znamená, že regióny (bunky) nemôžu splynúť
ani zaniknúť.
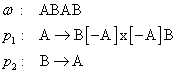
Obr.
5.9
Príklad mapového L-systému (Prusinkiewicz et al., 1990).
Rast rastliny ovplyvňujú vnútorné
(endogénne), ale aj vonkajšie (exogénne) faktory. Pôsobenie vnútorných faktorov
(prenos genetickej informácie, šírenie
hormónov, živín a vody) možno simulovať pomocou kontextových L-systémov.
Na simuláciu vplyvu vonkajších podmienok (vplyv svetla, prítomnosť
mechanických prekážok) bolo navrhnutých niekoľko techník, žiadna
z nich však neumožňovala kombinovať súčasne vplyv
endogénnych aj exogénnych faktorov. Pritom v prírode je takáto kombinácia
zjavná, napríklad pri odpovedi rastliny na prítomnosť alebo
neprítomnosť svetla, pri stretnutí sa s prekážkou, prerezávaní resp.
orezaní konárov (Obr.
5.10) alebo pri reakcii na napadnutie hmyzom.
Obr.
5.10
Odpoveď stromu na orezávanie (malé krúžky predstavujú spiace púčiky,
veľké naznačujú pozíciu šíreného signálu) a model záhrady
(Prusinkiewicz et al., 1994).
Ako riešenie tohto problému boli predstavené enviromentálne L-systémy (Prusinkiewicz
et al., 1994). Generovaný reťazec sa interpretuje po každom
derivačnom kroku a hodnoty korytnačky sa vrátia ako parametre
pre vyhradené moduly (query
modules) v reťazci. Každý derivačný krok prebieha ako
v parametrických L-systémoch s tým, že na rozdiel od nich,
v enviromentálnych L-systémoch zostávajú parametre spojené s modulmi
(query modules) nedefinované.
Počas grafickej interpretácie je týmto modulom priradená hodnota
v závislosti od pozície a orientácie korytnačky
v priestore. Parametre možno použiť ako argumenty používateľom
definovaných funkcií popisujúcich geometrický tvar, do akého sú stromy orezané.
Otvorené L-systémy (Prusinkiewicz et al., 2003)
rozširujú funkčnosť enviromentálnych L-systémov, a to použitím
symbolov pre komunikáciu medzi rastlinou a prostredím. Prostredie sa
nereprezentuje jednoduchou funkciou, ale stáva sa aktívnym procesom a môže
reagovať na informácie, prichádzajúce od rastliny. Rastliny sa preto
modelujú ako otvorené kybernetické systémy, ktoré odosielajú
a zároveň prijímajú informáciu z prostredia (Obr. 5.11). Odovzdávanie a príjem informácie
umožňujú tzv. komunikačné moduly (comunication modules).
Obr.
5.11
Simulácia interakcie rastliny s prostredím (Prusinkiewicz, 2000,
Prusinkiewicz et al., 2003).
Modely rastlín
založené na teórii L-systémov umožňujú (Prusinkiewicz, 2004a):
·
napodobniť stavbu a rast
rastlín,
·
ukázať vplyv rôznych
parametrov (napr. uhol vetvenia, veľkosť konárov, rýchlosť
predlžovania) na výsledný vzhľad rastlín,
·
simulovanie fyziologických
procesov v rastline, vplyvov človeka (orezávanie, vytváranie bonsaiov) a
vonkajších podmienok (vplyv svetla, vody, nedostatku miesta, prekážok) na jej
vývin,
·
simulovanie nielen samotných
rastlín, ale celých ekosystémov.
Hoci hlavné
uplatnenie L-systémov je pri modelovaní bylinných rastlín, boli použité aj na
modelovanie stromov. Prusinkiewicz (1990) ukázal, že pomocou L-systémov je
možné vygenerovať modely stromov, ktoré vytvorili Honda (1971) a Aono
a Kunii (1984). V osobitnej práci ukázal Shebell (1986), že pomocou
L-systémov sa dajú vygenerovať architektonické modely stromov, ktoré
popísali Hallé, Oldeman a Tomlinson (1978). L-systémy použil pri modelovaní
stromov tiež Shlyakhter (2001).
Okrem modelovania
rastlín boli L-systémy použité aj na simuláciu rastu tráviacej sústavy
u ľudských embryí (Ďurikovič et al., 1998), cestných sietí
v mestách (CityEngine, 2007), tvorbu hudby (Prusinkiewicz, 1986b,
Manousakis, 2006, LMUSe,
2007) ale aj na generovanie klasických lineárnych deterministických fraktálov
(Prusinkiewicz et al., 1990, Peitgen et al., 1992).
Pri modelovaní
stromov, ich stavby a funkcie, si treba stanoviť spôsob reprezentácie štruktúry
ich koruny, kmeňa, koreňov, teda ich architektúry. Pretože ciele
modelovania sa môžu líšiť podľa aplikácií, existuje široká škála
rôznych reprezentácii (Godin, 2000). Podľa neho možno stavbu rastliny
reprezentovať kombináciou troch informácií:
·
Dekompozícia (decomposition
information) – popisuje rôzne zložky (moduly), ktoré vytvárajú rastlinu.
·
Geometrická informácia (geometrical
information) – popisuje tvary a priestorové umiestnenie jednotlivých
zložiek. Zložky sa uvažujú nezávisle od seba.
·
Topologická informácia (topological
information) – popisuje vzájomný vzťah a spojenie medzi zložkami. Táto
informácia vyjadruje hierarchiu medzi zložkami v systéme rozkonárenia.
Podľa
stupňa komplexnosti reprezentácie rastliny, hovoríme o troch
reprezentáciách:
·
Globálne reprezentácie (global representations) – rastlina je
reprezentovaná ako celok. V tomto prípade nie je potrebné uvažovať
jednotlivé časti rastliny.
·
Modulárne reprezentácie (modular representations) – spoliehajú sa
na opis určitých častí rastliny (modulov), ktorých opakovaním je
vytvorená celá rastlina.
·
Viacúrovňové reprezentácie (multi-scale representations) – uvažujú
sa hierarchie modulov majúce rôznu veľkosť.
Pre našu potrebu
je zatiaľ dôležité podrobnejšie vysvetlenie modulárnej reprezentácie
rastlín, ktorú popíšeme v nasledujúcej časti.
Matematické modely v botanike zodpovedajú
rôznym úrovniam organizácie rastlín (..., molekula, organela, bunka, orgán,
rastlina, porast, ekosystém, bióm, ...) (Prusinkiewicz et al., 1996). Ak sa
zameriame na rastlinu, môžeme ju pokladať za súbor diskrétnych stavebných
jednotiek v priestore, modulov (modules), ktoré sa postupne vyvíjajú.
Väčšinou moduly reprezentujú opakujúce sa základné štrukturálne časti
rastliny, ako sú kvety, listy, články (internódiá), vrcholy alebo ich
zoskupenia, napríklad konáre alebo články stonky s priľahlým
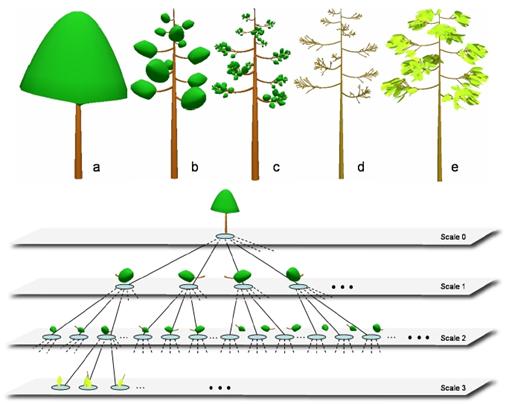
listom alebo pazušným púčikom (Obr. 6.1). Cieľom modelovania na modulárnej úrovni je
popísať vývoj rastliny ako celku a najmä vytváranie (emergence) rastlinného tvaru ako výsledku integrácie vývoja a pôsobenia
jednotlivých modulov (Prusinkiewicz et al., 2003).
Obr. 6.1
Príklady modulov a ich zoskupení (upravené podľa Prusinkiewicz et al.,
2003).
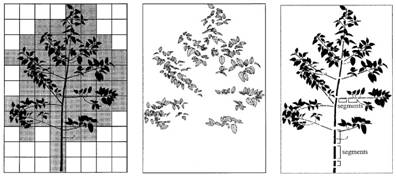
Pri modulárnej reprezentácii možno uvažovať dva
základné typy dekompozície, a to priestorovú
(spatial) a štrukturálnu (organ-based)
dekompozíciu (Godin, 2000). Pri
priestorovej dekompozícii je rozloženie modulov v priestore aproximované
rozdelením priestoru na bunky konštantnej veľkosti a označením
buniek, ktoré obsahujú moduly. Štrukturálna dekompozícia berie do úvahy
konkrétne moduly. Môžeme ju rozdeliť na dve triedy, geometrickú a topologickú
dekompozíciu. Pri geometrickej dekompozícii sa berie do úvahy len geometria
modulov a ich umiestnenie v priestore, kým pri topologickej
dekompozícii sa uvažujú aj spojenia medzi jednotlivými modulmi (Obr.
6.2).
Obr. 6.2 Modulárna reprezentácia rastliny, priestorová, geometrická a
topologická dekompozícia (zľava doprava) (Godin, 2000).
Vyššie spomínanú
klasifikáciu reprezentácie rastlín ďalej rozširuje Boudon (2006), ktorý
popisuje detailné, globálne a viacúrovňové reprezentácie stromov s dôrazom
na realistické a efektívne zobrazenie (rendering) (Obr. 6.3). Podobne ako v predchádzajúcej časti, zameriame sa na podrobnejší
popis detailnej reprezentácie rastlín.
Obr. 6.3 Reprezentácia stromov
(Boudon et al., 2006).
Detailné reprezentácie sa snažia zobraziť
geometriu stromu tak, aby vyzerala realisticky aj pre blízke pohľady.
Stromy majú dve odlišné časti, nerozkonárený kmeň a bohato
rozkonárenú korunu (ktorú vytvárajú konáre, na ktorých spravidla rastú listy),
čo si vyžaduje použitie rôznych techník na modelovanie a renderovanie
jednotlivých častí stromu. V snahe dosiahnuť realistické zobrazenie,
stromy sú chápané ako súbor modulov (listy, konáre a iné)
a geometrický model je vytvorený ich spojením.
Konáre sú obyčajne reprezentované množinou
pospájaných základných geometrických telies, na ktoré môže byť aplikovaná
textúra predstavujúca kôru. Pri návrhu realistickej reprezentácie kmeňa
a konárov je zložitou úlohou vytvorenie spojitého modelu z diskrétnej
množiny geometrických útvarov (geometrical
primitives). Preto je hlavným problémom definícia presnej reprezentácie miest, kde
sa spájajú, respektíve sa vetvia konáre (branch
junctions).
Najjednoduchšia reprezentácia konárov stromu je
pomocou valcov (3D cylinders) (Obr.
6.4). Vo svojich modeloch ich používa
(Kawaguchi, 1982, De Reffye et al., 1988, Prusinkiewicz et al., 1990).
Obr. 6.4 Geometria a príklad rozkonárovania s
použitím mnohostenov (Kawaguchi, 1982).
Bohužiaľ,
tieto techniky nezachytávajú správne geometriu tých miest, z ktorých
vychádzajú nové konáre a môžu sa objaviť rôzne medzery alebo prerušenia.
V snahe odstrániť tento problém bolo navrhnutých niekoľko
ďalších reprezentácií.
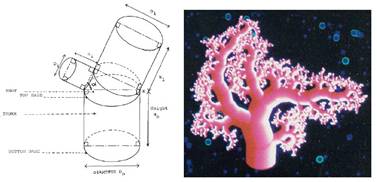
Max (1990) predstavil novú reprezentáciu pomocou útvaru, ktorý nazýva cone-sphere. Ten pozostáva z dvoch
gúľ a časti kužeľa alebo valca, ktorý leží medzi nimi
a dotýka sa ich (Obr.
6.5). Vonkajší povrch spojenia je tvorený
časťou gule, výsledkom čoho je plynulý prechod z jednej
časti na druhú. Na vnútornej strane sa však dotyčnice pretínajú skôr,
čo vedie k nespojitosti. Na zlepšenie výsledku je použitá zmiešavacia
metóda (blending method) založená na
jednoduchej lineárnej interpolácii. Táto reprezentácia umožňuje efektívne
modelovať jednotlivé konáre, ale prináša problémy pri použití textúr
v miestach spájania. V súvislosti s týmto problémom boli
navrhnuté ďalšie riešenia, formulované implicitne (Hart et
al., 1996, Galbraith et al., 2004).
Obr. 6.5 Cone-Sphere
(Max, 1990).
Na ďalšie
zlepšenie prispel Bloomenthal (1985), ktorý reprezentoval konáre pomocou
zovšeobecnených valcov (generalized
cylinders) (Obr. 7.2).
Bloomenthal tiež
predstavil implicitné povrchy (implicit
surfaces) (Bloomenthal, 1995). Na rozdiel od iných metód, implicitné
povrchy umožňujú vytvoriť ľubovoľné vetviace štruktúry, bez
ohľadu na ich komplexnosť. Ich výpočtová zložitosť je však
vysoká (Galbraith et al., 2004a, Galbraith et al., 2004b).
Obr. 6.6 Implicitná reprezentácia topoľa (Galbraith et al., 2004a).
Alternatívnu metódu na tvorbu hladkých povrchov
predstavujú subdivision surfaces (Obr. 6.7).
Obr. 6.7 Subdivision surfaces
(Tobler et al., 2002).
Prehľad techník použitých na textúrovanie kôry
a listov možno nájsť v (Boudon et al., 2006).
V literatúre sa stretávame s viacerými
klasifikáciami, ktoré sa snažia utriediť rôzne metódy navrhnuté pre
modelovanie rastlín, konkrétne stromov a zovšeobecniť výhody a nevýhody
jednotlivých prístupov:
1) Rozdelenie metód na biologicky motivované
(biology-based) a metódy vychádzajúce
z geometrie (geometry-based)
(Shlyakhter, 2001):
2) Biology-based – tieto metódy v sebe zahŕňajú
biologické zákonitosti týkajúce sa rastu stromov, generujú vierohodné modely,
používateľ má však len veľmi malú kontrolu nad výsledkom, hlavne
výsledným tvarom stromu.
3) Geometry-based – tieto metódy ponúkajú väčšiu
kontrolu výsledného tvaru, ale modely nedosahujú botanickú vernosť.
4)
Quan
(2006) rozširuje predchádzajúce rozdelenie na metódy založené na pravidlách (rule-based
methods) alebo obrazoch (image-based methods):
5) Rule-based methods – používajú pevne stanovené pravidlá
alebo gramatiky:
6) L-systémy (Prusinkiewicz et al., 1990,
atď.).
7) Geometrické pravidlá (Weber et al., 1995).
8) Biologické pravidlá (De Reffye et al.,
1988, atď.).
Hoci tieto metódy umožňujú vytvoriť presvedčivé modely
stromov alebo celých ekosystémov, pravidlá alebo parametre, z ktorých vychádzajú
sú ťažko použiteľné pre laika.
9) Image-based
methods – umožňujú
vymodelovať konkrétny strom na základe jeho fotografií. Použité sú rôzne
prístupy:
10) Visual
hull (Sakaguchi et al.,
1999, Shlyakhter et al., 2001).
11) Objemová reprezentácia (Reche et al., 2004).
12) Structure
from motion (Quan et al.,
2006).
Nevýhodou pri týchto prístupoch je len približný tvar stromu s
nerealistickou štruktúrou konárov a veľkým počtom polygónov
(Shlyakhter et al., 2001) alebo prístup, ktorý neumožňuje získať
explicitnú geometriu konárov a listov a výsledný model nemožno ďalej
editovať (Reche et al., 2004).
13)
Rozdelenie
procedurálnych modelov na dve triedy – local-to-global
a global-to-local modely
(Prusinkiewicz et al., 2001, Boudon et al., 2003):
14) Modely local-to-global
– používateľ si určí jednotlivé časti (moduly) rastliny, ktoré
sa na základe použitého algoritmu spoja do komlexnej štruktúry. Kvôli
emergentnému charakteru rastlín býva však ťažké kontrolovať výsledný
tvar. Výnimkou sú prístupy, ktoré umožňujú kontrolovať tvar rastliny,
vznikajúci ako odpoveď na orezávanie (Prusinkiewicz et al., 1994).
15) Modely global-to-local
– na rozdiel od predchádzajúcich, používateľ charakterizuje celkový
vzhľad rastliny ako napr. jej siluetu alebo hustotu rozloženia konárov
(Reeves et al., 1985, Weber et al., 1995, Ďurikovič, 2003).
Algoritmus z tejto informácie odvodí detailnejšiu štruktúru rastliny. Tento
prístup ponúka priamejšiu a intuitívnejšiu kontrolu výsledného tvaru, a preto
sa uprednostňuje v aplikáciách, kde je dôležitá najmä vizuálna
stránka. Tieto aplikácie zahŕňajú aj odvodenie tvaru stromu z
fotografií (Shlyakhter et al., 2001, Quan et al., 2006) a interaktívnu kontrolu
tvaru (Prusinkiewicz et al., 2001, Boudon et al., 2003, Lintermann et al.,
1999, Deussen et al., 2005). Výhodou týchto prístupov je, že parametre použité
pre tvorbu detailnejšej stavby sú odvodené (zdedené) z parametrov popisujúcich
celkovú stavbu stromu. Nevýhodou pri interaktívnom modelovaní je to, že ide o
zdĺhavý proces a ľahko sa môže stať, že používateľ sa zamotá,
ak si stanoví veľký počet parametrov a stráca tak možnosť
intuitívnej kontroly nad modelom. Tento problém sa však dá vyriešiť
použitím viacúrovňovej reprezentácie rastlín (multiresolution editing) (Boudon et al., 2003).
Simulácia vetviacich štruktúr má
v počítačovej grafike relatívne dlhú históriu. Prvý model, ktorý
bol navrhnutý, vychádza z koncepcie celulárnych automatov (cellular automata) (Ulam, 1966). Vetviaci vzor (branching pattern) sa vytvára iteratívne, začínajúc jedinou farebnou bunkou v
trojuholníkovej mriežke. Postupne sa farbia bunky, ktoré sa dotýkajú práve
jedného vrchola bunky, ktorá bola zafarbená v predchádzajúcom
iteračnom kroku.
Pre tento prístup (a jeho ďalšie rozšírenia) je
charakteristický dôraz na interakciu medzi rôznymi časťami rastúcej
štruktúry a rovnako štruktúry a prostredia. Hoci interakcie
ovplyvňujú vývoj rastlín, pridávajú tiež na zložitosti modelov. To
vysvetľuje, prečo jednoduché modely, ktoré ignorujú aj základné
faktory, napríklad kolíziu medzi konármi, prevládajú dodnes.
Obr. 7.1 Špecifikácia
geometrie stromu podľa Hondu (1971).
Prvý jednoduchý model stromu navrhol Honda (1971),
ktorý študoval tvary stromov s použitím nasledujúcich pravidiel (Obr. 7.1):
·
Segmenty stromu sú rovné, ich
hrúbka (obvod) sa neuvažuje.
·
Materský segment vytvorí
počas jedného rozkonárovania (branching process) dva dcérske
segmenty.
·
Dĺžky dcérskych segmentov sa
skrátia konštantnými škálovacími koeficientmi 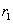 a
a 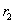 , vzhľadom na materský segment.
, vzhľadom na materský segment.
·
Materský segment a jeho dva
dcérske segmenty ležia v jednej rovine (branch plane). Dcérske segmenty vytvárajú vzhľadom na materský
segment konštantné uhly (branching angles)
 a
a 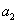 .
.
·
Rovina rozkonárenia je
vzhľadom na gravitáciu pevne stanovená, a to tak, aby bola čo najbližšie
k horizontálnej rovine. Výnimku tvoria vetvy, ktoré vyrastajú
z hlavného kmeňa. V ich prípade sa zachová konštantný uhol odchýlky (divergence angle) medzi za sebou
vychádzajúcimi bočnými segmentmi.
Zmenou parametrov
získal širokú škálu tvarov, podobajúcich sa na stromy a vďaka
ďalším vylepšeniam sa jeho model používal na skúmanie spôsobov
rozkonárovania u reálnych stromov.
Niekoľko
ďalších rozšírení založených na jeho výsledkoch navrhli Aono a Kunii
(1984). Zároveň boli prvými autormi, ktorý skúmali použitie L-systémov na
tvorbu botanických stromov. Vychádzali z pôvodnej definície L-systémov,
pričom zistili, že je nevhodná na modelovanie rozkonárených štruktúr
u vyšších rastlín.
Doteraz spomínané modely reprezentovali „kostru
stromu“ s použitím rovných čiar konštantnej alebo meniacej sa hrúbky.
Podstatné realistické vylepšenie modelov dosiahli Bloomenthal (1985)
a Oppenheimer (1986), ktorí predstavili oblé konáre, dôkladne namodelované
povrchy v miestach rozkonárenia a otextúrovanú kôru a listy.
Bloomenthal sa namiesto štúdia rôznych typov
rozkonárovania zameral na zlepšenie realizmu výsledného modelu (Obr. 7.2). Stromy reprezentoval ako množinu bodov v priestore
a ich spojení (konárov). Konáre sú modelované ako "zovšeobecnené valce" (generalized cylinders), ktoré sú reprezentované ako priestorové krivky
interpolujúce body a postupnosť obrysov (cross sectional contour)
kolmých na krivku. Povrch stromu sa teda znázorňuje ako zovšeobecnený
valec s pospájanými kruhovitými (konáre) alebo zakrivenými (kmeň) obrysmi
rôzneho polomeru.
Obr. 7.2 Špecifikácia geometrie stromu podľa
Bloomenthala (Bloomenthal, 1985).
Oppenheimer zaviedol fraktálny model stromu, ktorého
geometria a topológia je kontrolovaná číselnými parametrami analogickými
ku DNA živých organizmov. Zmenou genetickej informácie (parametrov) nastáva
zmena geometrie objektu, výsledný model však ostáva samopodobný, hoci menením
parametrov to nemusí byť až také viditeľné (Obr. 7.3).
Obr. 7.3 Fraktálny
model stromu a stonky a listov maliny (Oppenheimer, 1986).
Reeves a Blau (1985) sa zamerali na tvorbu
lesných scén a použitím stochastického prístupu, systému častíc (particle
systems), vytvárali veľmi komplexné obrázky, ktoré ale nerešpektovali
biologické zákonitosti (Obr. 7.4).
Obr. 7.4 Lesná scéna vytvorená pomocou systému
častíc (Reeves et al., 1985).
De Reffye (1988)
navrhol model stromu, ktorý vychádza z biologických zákonitostí (ako stromy
rastú, obsadzujú priestor, kde a ako vyrastajú listy, kvety alebo plody).
Nezameriava sa na geometriu stromu ako Bloomenthal, ale zdôrazňuje skôr
topologický model stromu. Hlavná myšlienka spočíva v modelovaní aktivity
púčikov v diskrétnych časových intervaloch. Na základe signálu
môže púčik:
·
stať sa kvetom a
odumrieť (a zaniknúť),
·
zmeniť sa na spiaci
púčik,
·
predĺžiť sa na
článok (internódium), ktorý bude ukončený novým vrcholovým
púčikom a jedným alebo viacerými bočnými púčikmi, ktoré sa
objavia v pazuchách listov.
Tieto zmeny sa dejú na základe stochastických
zákonov charakteristických pre jednotlivé druhy a variety. Podľa
stochastických pravidiel sú tiež vypočítané geometrické parametre, ako
napríklad dĺžka a priemer článkov alebo uhol vetvenia. Všetky
pravdepodobnostné zákonitosti môžu byť merané experimentálne, napríklad na
poľnohospodárske ciele, a merania môžu potvrdiť správnosť
modelu. Zmenou pravdepodobností možno získať architektonické modely
drevín, ktoré popísali Hallé, Oldeman a Tomlinson (1978).
Sila takéhoto prístupu spočíva v tom, že
pomocou tých istých procedurálnych metód možno vytvoriť rôzne
modely rastlín, od drevín ako
napríklad jedľa, borovica, čerešňa, vŕba, topole až po
bylinné druhy (Obr. 7.5).
Obr. 7.5 Ukážky
modelov stromov vytvorených podľa (De Reffye et al., 1988).
Obr. 7.6 Ukážky
modelov stromov vytvorených podľa Webera (1995).
Weber a Penn
(1995) sa namiesto rešpektovania biologických zákonitostí pri modelovaní
stromov zamerali na celkový geometrický tvar. Ich model je založený na dvoch
častiach, stonke (kmeň a konáre) a liste. Kmeň stromu pozostáva
zo zaoblených kužeľovitých častí, ktoré sa môžu na rôznych miestach
rozdeliť a vytvoriť nové valcovité štruktúry, ktoré sa môžu
ďalej deliť (rozkonárovať). Mnohé atribúty nových konárov (napr.
dĺžka) sú odvodené od rodičovských, môžu však byť aj úplne iné.
Použitím malého počtu parametrov vytvorili len na základe pozorovania a
porovnávania tvarov reálnych stromov, niekoľko rôznych modelov stromov
podobných reálnym druhom (Obr. 7.6). Autori tiež predstavili viacúrovňovú
reprezentáciu stromov, umožňujúcu efektívne zobrazovanie väčšieho
počtu stromov v scéne.
Viacerí autori
publikovali rôzne procedurálne metódy umožňujúce generovať stromy,
Sakaguchi (Sakaguchi et al., 1999) sa však zameral na inverzný problém, a to
ako odvodiť tieto metódy pre konkrétny strom, na základe jeho fotografií.
Takáto potreba môže vzniknúť ak chceme vytvoriť model nejakého
konkrétneho stromu (napr. historicky alebo kultúrne významného), nachádzajúceho
sa v scéne. Hľadať parametre, z ktorých by sme mohli odvodiť
tvar daného stromu je príliš zdĺhavé a intuitívne a môže spôsobiť
veľa neželaných efektov.
Tento prístup
vylepšil Shlyakhter (Shlyakhter et al., 2001), ktorý odvodzuje hlavnú štruktúru
(kostru) stromu priamo z jeho celkového tvaru. Vychádza z fotografií stromu,
štyroch až pätnástich, ktoré zachytávajú najmenej 135° okolo stromu. Pozícia a
orientácia fotoaparátu pre každý obrázok je známa. Proces rekonštrukcie
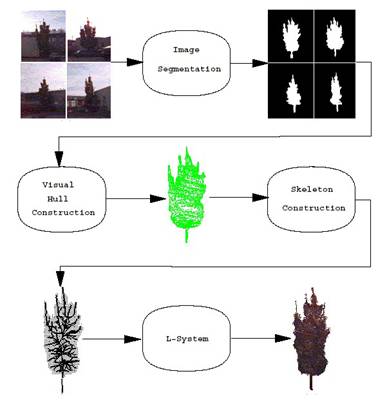
pozostáva zo štyroch krokov (Obr. 7.8):
1) Segmentácia obrázkov − všetky
obrázky sú segmentované na strom a pozadie. Tento proces je manuálny, v
grafickom editore sa naznačí obrys, silueta stromu.
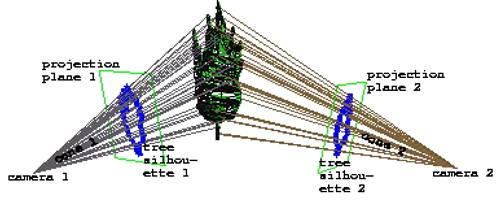
2) Visual
hull − každá
silueta získaná zo vstupných fotografií sa aproximuje mnohouholníkom.
Projekciou jeho obrysu vzniká „kužeľ“ (n-boký nekonvexný ihlan), ktorý
ohraničuje 3D strom ležiaci v ňom, takže vypočítaním
prieniku všetkých kužeľov, vznikne aproximácia trojrozmerného tvaru
stromu, visual hull (Laurentini,
1994).
Obr. 7.7 Rekonštrukcia tvaru stromu zo siluet
(Shlyakhter et al., 2001).
3) Vytvorenie kostry stromu − kostra
stromu (kmeň a niekoľko ďalších stupňov rozkonárenia) je
hľadaná ako aproximácia osi (medial
axis) obalu (visual hull). Autori
vychádzajú z myšlienky, že stromy sa v snahe prijať čo najviac
slnečnej energie potrebnej pre fotosyntézu, snažia rozložiť konáre v
priestore čo najefektívnejšie. Tieto pozorovania naznačujú, že kostru
stromu možno prirovnať ku osi jeho tvaru, ktorá predstavuje v istom zmysle
optimálnu kostru.
4) L-systém − vytvorená kostra stromu
sa použije ako axióma pre otvorený L-systém, ktorý pridá posledným dvom
stupňom konárov púčiky, ktoré umožnia rast ďaších konárov a
listov. L-systém zároveň zabezpečí rozloženie konárov a listov a tiež
to, aby všetky konáre, ktoré prerastú tvar stromu, boli odrezané.
Obr. 7.8 Proces rekonštrukcie 3D modelu stromu z
fotografií (Shlyakhter et al., 2001).
Obr. 7.9 Časová náročnosť
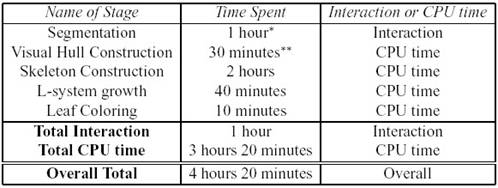
jednotlivých krokov rekonštrukcie (Shlyakhter et al., 2001).
Veľkou
nevýhodou tejto metódy je okrem značnej prácnosti a vysokého počtu
polygónov, len približná kostra stromu, ktorá nie je obrazom reálneho
rozkonárenia (Obr. 7.10).
Obr. 7.10 Ukážky kostry stromu a porovnanie
fotografie reálneho stromu s vytvoreným modelom (zľava doprava)
(Shlyakhter et al., 2001).
Poloautomatickú, interaktívnu techniku
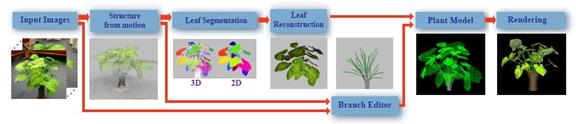
na modelovanie rastlín priamo z fotografií navrhol Quan (2006). Výsledný model
je geometrický a môžno s ním ľahko manipulovať, prípadne ho
ďalej editovať.
Obr. 7.11 Proces rekonštrukcie modelu z fotografií
(Quan et al., 2006).
Ich metóda pozostáva z troch krokov (Obr.
7.11):
1) Získanie obrazov (fotografií) a použitie
algoritmu structure from motion – vstupné dáta predstavuje 30 až 45
vzájomne sa prekrývajúcich fotografií, voľne vzatých dookola rastliny, tak
aby bola rastlina zachytená z viacerých pohľadov. Fotoaparát nemusí
byť kalibrovaný, malo by sa však vyhnúť zmenám expozície v dôsledku
nerovnakého osvetlenia alebo tieňov. Ďalej sa použije structure
from motion algoritmus (Lhuiller et al., 2005), vďaka ktorému sa
získajú parametre fotoaparátu a oblak 3D bodov. Tieto body pomôžu pri
určení tvaru rastliny. Každý bod je spojený s informáciou o tom, z ktorej
fotografie pochádza, čo má význam pri ďalších krokoch, a to
segmentácii listov a konárov.
2)
Segmentácia
a rekonštrukcia listov – pomocou navrhnutej graph-based optimalizačnej
techniky sa 3D body a pixely fotografií súčasne rozdelia do diskrétnych
množín, kde každá množina reprezentuje jeden list. Táto časť procesu
segmentácie prebieha automaticky, ale v ďalšom kroku môže do nej vstúpiť
aj používateľ, ktorý môže interaktívnym spôsobom zefektívniť a
vylepšiť proces segmentácie. Po každej zmene sa systém hneď
aktualizuje. Keďže listy rastliny sú prevažne rovnaké, na rekonštrukciu
listov si používateľ vyberie jeden vzorový list, ktorý je ďalej
upravený a použitý namiesto ostatných listov, čo zvyšuje robustnosť
systému (Obr. 7.12).
3)
Interaktívna
rekonštrukcia konárov – kým listy sú rekonštruované prevažne automaticky, pri konároch
je to kvôli ich komplexnosti problém. Autori považujú rekonštrukciu konárov
pomocou L-systémov za zložitú, pretože používateľ nemusí vedieť, aké
vstupné parametre použiť, aby dostal výsledný tvar. Namiesto použitia
L-systémov, navrhli editor, ktorý umožňuje interaktívnym spôsobom
kontrolovať výsledný tvar konárov. Konáre sú modelované ako zovšeobecnené
valce (generalized cylinders), ktorých os tvorí priestorová krivka.
Samotný proces rekonštrukcie pozostáva z niekoľkých krokov.
Používateľ najskôr kreslí krivky, ktoré reprezentujú jednotlivé konáre a
následne ich na základe vstupných fotografií pre ľubovoľné
pohľady upravuje, mení ich umiestenie v priestore, tvar, polomer alebo
určuje, či na konci konára budú rásť listy (Obr. 7.12). Výsledný model vznikne spojením
rekonštruovaných konárov a listov, ktoré sú automaticky pripojené k najbližším
konárom.
Obr. 7.12 Segmentácia a
rekonštrukcia listov a konárov (zľava doprava) (Quan et al., 2006).
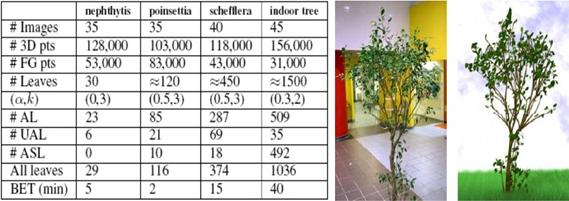
Metóda bola
testovaná na izbových rastlinách, kde ponúkala veľmi dobré výsledky,
vďaka malému počtu konárov a veľkým listom. Bola vyskúšaná aj na
modelovanie menšieho (izbového) stromu, v tomto prípade sa však kvôli
väčšej komplexnosti objektu (väčší počet konárov aj listov,
ktoré sú navyše menšie, v porovnaní s izbovými rastlinami) zvýšila
výpočtová zložitosť a výsledok bol menej presný (Obr. 7.13).
Obr. 7.13 Štatistika rekonštrukcie a porovnanie
fotografie stromu s modelom. FG – foregrounds, AL – automatic leaves, UAL
– user assisted leaves, ASL – additional synthetic leaves, BET – branch edit
time (Quan et al., 2006).
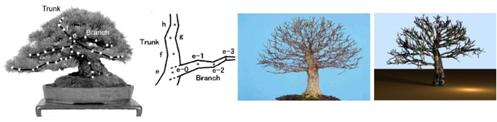
Ďurikovič (2003) predstavil
metódu na modelovanie presného tvaru stromov, ktorú ukázal na modelovaní
bonsaiov, u ktorých je podstatný práve ich vonkajší tvar. Tvar je odvodený z
fotografií stromu a nameraných dát. Na základe nameraných dát pre vybrané
konáre odvodí funkciu aproximujúcu dáta, ktoré sú použité na vytvorenie
L-systému, schopného reagovať na vonkajšie vplyvy prostredia,
prijímať a zároveň odovzdávať informácie pomocou
komunikačných modulov. L-systém vygeneruje kostru stromu (hlavné konáre
vytvárajúce tvar stromu), majúcu rovnaký tvar ako odvodená funkcia. Pomocou
L-systému sú generované aj drobné konáre (konáre vyššieho stupňa) a listy.
Tie sú rozmiestnené okolo hlavných konárov podľa funkcie určujúcej
hustotu rozloženia. Konáre sú reprezentované ako zrezané kužele vychádzajúce z
uzlov, s výškou odvodenou od ich začiatočného a koncového polomeru.
Polomer zodpovedá aktuálnej hrúbke konáru danej jeho umiestnením. Kým parametre
hlavných konárov sú dané z nameraných dát, parametre menších konárov sú
vypočítané náhodne z priemerných hodnôt. Listy sú reprezentované ako
trojuholníková sieť a ihlice ako dlhé úzke kužele.
Obr. 7.14 Znázornenie meraných
bodov a porovnanie fotografie s vygenerovaným stromom (Ďurikovič,
2003).
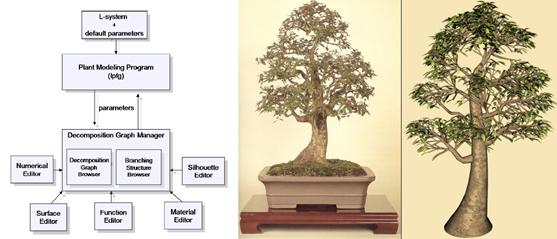
Metódu na kontrolovanie veľkého
množstva vstupných parametrov potrebných pri detailnejšom popise a modelovaní
rastlín navrhol (Boudon et al., 2003) a ilustroval ju na interaktívnom
modelovaní bonsaiov. Ide o viacúrovňovú reprezentáciu, pri ktorej popisujú
štruktúru rastlín pomocou decomposition graph. Je to strom (v
teoretickom zmysle), ktorý zachytáva hierarchickú štruktúru rastliny, na
základe stupňa rozkonárenia (Obr. 7.15). Uzly tohto grafu obsahujú parametre, ktoré popisujú časti
stromu na jednotlivých hierarchických úrovniach, a teda na rôznych
stupňoch detailu. V procese interaktívneho modelovania stromu sú parametre
popisujúce vyššie stupne konárov zdedené z parametrov popisujúcich strom ako
celok. Používateľ môže manipulovať s týmito parametrami alebo ich
editovať bez ohľadu na dedičnosť. Model je vytváraný
algoritmicky, s parametrami uloženými v decomposition graph. Na
začiatku modelovacieho procesu si používateľ špecifikuje algoritmus
(L-systém), ktorý vytvorí decomposition graph a definuje parametre pre niektoré
uzly. Pre tie uzly, ktoré nemali explicitne definované parametre, sú parametre
zdedené. Následne môžu byť parametre interaktívne editované. Pri každej
zmene sa algoritmus znovu spustí a výsledný tvar stromu sa aktualizuje (Obr.
7.16).
Obr. 7.15 Viacúrovňová reprezentácia štruktúry
stromu (hore) a príslušný decomposition
graph (dole) (Boudon et al., 2003).
Obr. 7.16 Interakcia rôznych komponentov počas
procesu modelovania (vľavo), porovnanie fotografie s modelom (vpravo)
(Boudon et al., 2003).
Modelovanie
rastlín má medzivedný charakter a odráža diverzitu existujúcich
a možných aplikácií modelov (Prusinkiewicz, 2004a).
·
Vedecké využitie:
·
Biológia – výskum hypotéz
týkajúcich sa genetických, fyziologických alebo biomechanických zákonitostí
rastu rastlín alebo biologických štruktúr. Klasifikácia architektúry (vonkajšej
stavby, tvaru) rastlín.
·
Ekológia – skúmanie vzťahov
medzi samotnými rastlinami a tiež rastlinami a prostredím.
·
Evolúcia – analýza tvaru rastlín z
pohľadu evolúcie.
·
Praktické využitie, kde je
podstatná architektúra rastlín:
·
Šľachtenie rastlín a
genetické inžinierstvo – špecifikácia "cieľovej" rastliny na
základe určenia stavby tela optimálnej pre príjem slnečného žiarenia,
úrodnosť, odolnosť voči poškodeniu, kvalitu a kvantitu plodov,
kvetov, vzhľadu, atď.
·
Ošetrovanie rastlín – určenie
stratégii rastlín pri obsadzovaní priestoru a optimálne ošetrovanie
(orezávanie, použitie chemikálií, stanovenie vhodnej teploty) v ovocných
sadoch, skleníkoch, záhradách alebo v bytoch.
·
Krajinná architektúra – simulácia
interakcií medzi stromami a rôznymi stavbami, napr. spôsob odstraňovania
konárov, aby sa zabránilo kolízii s elektrickým vedením.
·
Praktické využitie, kde nie je
podstatná architektúra rastlín, ale jej zahrnutie môže vylepšiť hodnotu
predpovedných modelov:
·
Simulovanie prítomnosti a šírenia
patogénov u rastlín.
·
Biologická kontrola prítomnosti
burín (inváznych druhov) – skúmanie ich stavby a vplyvu rôznych patogénov alebo
prítomnosti bylinožravcov na ich výskyt.
·
Simulovanie vplyvu pesticídov.
·
Aplikácie vo vyučovaní:
·
Vysvetlenie a ilustrácia
procesov prebiehajúcich v rastlinách, vzťahov medzi rastlinou
a prostredím. Využitie vo vyučovacom procese, pri cvičeniach.
Virtuálne modely
rastlín prispievajú k pochopeniu rôznych mechanizmov prebiehajúcich v
rastlinách počas ich rastu. Napriek tomu je ich využitie v praxi (napr. v
záhradníctve, poľnohospodárstve, lesníctve) zatiaľ nevýrazné.
Prusinkiewicz (2004a) naznačuje niekoľko faktorov, ktoré môžu
byť príčinou tohto stavu:
·
Fyziologické mechanizmy
ovplyvňujúce funkciu rastlín nie sú natoľko známe, aby mohli
poskytnúť pevný základ pre tvorbu prognostických modelov. Ešte menej je
známe o genetickej podstate vývinu rastlinných foriem.
·
Metódy na modelovanie rastlín a
tvorba príslušného softvéru patria stále do oblasti aktívneho výskumu a bude
chvíľu trvať kým sa začnú používať na riešenie praktických
problémov.
·
Tvorba rastlinných modelov je
úsilím viacerých vedných odborov. Kombinuje v sebe rôzne aspekty biológie,
matematiky, počítačovej grafiky a špecifických modelovacích
zručností (umenia), preto je ťažké nájsť ľudí alebo
vytvoriť tímy, ktoré by mali žiadané odborné znalosti.
·
Tvorba presných modelov vyžaduje
rozsiahle merania rastlín, a preto je drahá.
Otvoreným
problémom tiež zostáva začlenenie genetických mechanizmov do virtuálnych
rastlín, čo by mohlo priniesť nový pohľad na pochopenie
vývinových procesov u rastlín.
V práci sme
predstavili prehľad metód na modelovanie stromov. Existuje viacero
programov, ktoré umožňujú viac či menej realistické modelovanie
stromov. Medzi najznámejšie patrí XFrog, SpeedTree, vlab/L-studio. Prehľad
jednotlivých programov spolu s požiadavkami na systém, výhodami a nevýhodami
možno nájsť na stránke Virtual Terrain Project (Plant Generation Software
Packages, 2007).
Zoznam bibliografických
odkazov
ABELSON, H.,
DISESSA, A.A. 1982. Turtle geometry.
Cambridge : M:I:T: Press, 1982. Zroj: PRUSINKIEWICZ, P., LINDENMAYER, A.
The algorithmic beauty of plants. New York : Springer Verlag, 1990
Adobe
Photoshop [online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.adobe.com/products/photoshop/photoshop/>.
AKENINE-MÖLLER,
T., HAINES, E., 2002. Real-time rendering.
2nd ed. AK Peters, 2002.
AONO, M.,
KUNII, T.L. 1984. Botanical tree image generation. IEEE Computer Graphics and Applications. 1984, vol. 4, no. 5, p.
10-34. Zroj: PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic
beauty of plants. New York : Springer Verlag, 1990
BLOOMENTHAL,
J. 1985. Modeling the mighty maple. In SIGGRAPH ’85: Proceedings of the 12th
annual conference on Computer graphics and interactive techniques, 1985, p.
305-311.
BLOOMENTHAL,
J. 1995. Skeletal design of natural forms.
PhD thesis. University of Calgary, 1995. Zdroj:
BOUDON, F., MEYER, A., GODIN, Ch. 2006. A survey of computer representations of
trees for realistic and efficient rendering. Report de recherche. In LIRIS, 2006.
BOBÁK, M. et
al. 1992. Botanika : Anatómia
a morfológia rastlín. Bratislava : SPN, 1992.
BOUDON, F.,
PRUSINKIEWICZ, P., FEDERL, P. et al. 2003. Interactive design of bonsai trees.
In Computer Graphics Forum. Proceedings
of Eurographics, 2003, vol. 22, no. 3, p. 591-599.
BOUDON, F.,
MEYER, A., GODIN, Ch. 2006. A survey of computer representations of trees for
realistic and efficient rendering. Report de recherche. In LIRIS, 2006.
CityEngine
[online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.vision.ee.ethz.ch/~pmueller/wiki/CityEngine/Documents>.
COOMBES, A.J.
2001. Stromy. 2. vyd. Martin :
Osveta, 2001. ISBN 80-8063-066-6.
DE REFFYE, P.,
EDELIN, C., FRANÇON, J. 1988. Plant models faithful to botanical structure and
development. In SIGGRAPH ’88: Proceedings of the 15th annual conference on
Computer graphics and interactive techniques, 1988, p. 151-158.
DEUSSEN, O.,
LINTERMANN, B. 1997. A modelling method and user interface for creating
plants. In Proceedings of the conference
on Graphics Interface ’97, 1997, p. 189-197.
DEUSSEN, O.,
LINTERMANN, B. 2005. Digital Design of
Nature : Computer Generated Plants and Organics. Berlin : Springer-Verlag, 2005. ISBN 3-540-40591-7.
ĎURIKOVIČ, R., KANEDA, K., YAMASHITA, H. 1998. Animation of
biological organ growth based on L-systems. In Computer Graphics Forum,
1998, vol. 17, no. 3, p. 1-13.
ĎURIKOVIČ, R. 2003. Towards visual modelling of bonsai trees.
In Proceedings of the IASTED Visualization, Imaging, Image Processing –
VIIP2003, 2003, p. 797-802.
EBERT, D.S.,
MUSGRAVE, F.K., PEACHEY, D. et al. 1998. Texturing and modeling : A
procedural approach. 2nd ed. USA : Academic Press, 1998.
Environmental
Information Laboratory [online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.h.chiba-u.jp/terra/envinf4.htm>.
FABRIKA, M.,
Ďurský, J. 2005. Stromové rastové
simulátory. Zvolen : EFRA – Vedecká agentúra pre ekológiu a lesníctvo,
2005. ISBN 80-969434-7-2.
FERRARO, P.,
GODIN, Ch., PRUSINKIEWICZ, P. 2005. Toward a quantification of self-similarity
in plants. In Fractals, 2005, vol. 13, no. 2, p. 91-109.
GALBRAITH, C.,
MACMURCHY, P., WYVILL, B. 2004a. BlobTree Trees. In CGI ’04: Proceedings of
the Computer Graphics Interactional (CGI ’04), 2004, p. 78-85.
GALBRAITH, C.,
MÜNDERMANN, L., WYVILL, B. 2004b. Implicit visualization and inverse modeling
of growing trees. In Computer Graphics Forum (Proceedings of Eurographics
2004), 2004, vol. 23, no. 3, p. 351-360.
GODIN, Ch.
2000. Representing and encoding plant architecture: a review. In Annals of
Forest Science, 2000, vol. 57, no. 05-juin, p. 413-438.
Growth model
SIBYLA [online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.uni-forst.gwdg.de/~mfabrik/>.
HALLÉ, F.,
OLDEMAN, R.A.A., TOMLINSON, P.B. 1978. Tropical trees and forests: An
architectural analysis. Berlin : Springer Verlag, 1978. Zroj: PRUSINKIEWICZ,
P., LINDENMAYER, A. The algorithmic beauty of plants. New York :
Springer Verlag, 1990.
HART, J.C.,
BAKER, B. 1996. Implicit modeling of tree surfaces. In Implicit
surfaces ’96, 1996, p. 143-152.
HIEKE, K.
1978. Praktická dendrologie (1, 2). Praha : SZN, 1978.
HONDA, H.
1971. Description of the form of trees by the parameters of the tree-like body:
Effects of the branching angle and the branch length on the shape of the
tree-like body. In Journal of Theoretical Biology, 1971, 31, p. 331-338.
Zroj: PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic beauty of
plants. New York : Springer Verlag, 1990.
KAWAGUCHI, Y.
1982. A morphological study of the form of nature. In SIGGRAPH ’82:
Proceedings of the 9th annual conference on Computer graphics and interactive
techniques, 1982, p. 223-232.
KURTH, W.
1994. Growth Grammar Interpreter GROGRA
2.4 : A software tool for the 3-dimensional interpretation of stochastic,
sensitive growth grammars in the context of plant modelling. Introduction and
Reference Manual. Forschungszentrum Waldökosysteme der Universität
Göttingen, Göttingen, 1994.
LAURENTINI, A.
1994. The visual hull concept for silhouette-based image understanding. In IEEE Trans. Pattern Anal. Mach. Intell,
1994, vol. 16, no. 2, p. 150-162.
LHUILLIER, M.,
QUAN, L. 2005. A quasi-dense approach to surface reconstruction from
uncalibrated images. In IEEE Trans. Pattern
Anal. Mach. Intell., 2005, vol. 27, no. 3, p. 418-433.
LINTERMANN,
B., DEUSSEN, O. 1999. Interactive modeling of plants. In IEEE Comput. Graph. Appl., 1999, vol. 19, no. 1, p. 56-65.
LMUSe [online]
[cit. 2007-04-12]. Dostupné na internete: <http://www.geocities.com/Athens/Academy/8764/lmuse/lmuse.html>.
MALÁ, B.,
MLADÝ, L., MALÝ, M. 2000. Generálna
rekonštrukcia areálu zámku Budatín. Žilina : MGM, 2000.
MANDELBROT,
B.B. 1982. The fractal geometry of
nature. San Francisco : W. H. Freeman, 1982. Zdroj: PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic beauty
of plants. New York : Springer Verlag, 1990.
Mandelbrot set
[online] [cit. 2007-04-12]. Dostupné na internete: <http://en.wikipedia.org/wiki/Mandelbrot_set>.
MANOUSAKIS, S.
2006. Musical L-systems. Master’s
thesis. The Royal Conservatory, The Hague, 2006.
MAX, N. 1990.
Cone-spheres. In SIGGRAPH ’90: Proceedings of the 17th annual conference on
Computer graphics and interactive techniques, 1990, p. 59-62.
MĔCH, R.,
PRUSINKIEWICZ, P., HANAN, J. [online]. Extensions to the graphical
interpretation of L-systems based on turtle geometry [cit. 2007-04-12].
Dostupné na internete: <http://algorithmicbotany.org/lstudio/graph.pdf>.
NAKAMURA, A.,
LINDENMAYER, A., AIZAWA, K. 1986. Some systems for map generation. In
ROZENBERG, G., SALOMAA, A. (eds.) The Book of L. Berlin :
Springer-Verlag, 1997, p. 323-332. Zdroj:
PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic beauty of plants.
New York : Springer Verlag, 1990.
OPPENHEIMER,
P.E. 1986. Real time design and animation of fractal plants and trees. In SIGGRAPH
Comput. Graph., 1986, vol. 20, no. 4, p. 55-64.
PEITGEN,
H.-O., JÜRGEN, H., SAUPE, D. 1992. Chaos and fractals : New frontiers of
science. New York : Springer Verlag, 1992. ISBN 0-387-97903-4.
Plant
Generation Software Packages [online]. [cit. 2007-04-12]. Dostupné na
internete: <http://www.vterrain.org/Plants/plantsw.html>.
PRUSINKIEWICZ,
P. 1986a. Graphical applications of L-systems. In Proceedings on Graphics Interface ’86 / Vision Interface ’86, 1986,
p. 247-253.
PRUSINKIEWICZ,
P. 1986b. Score generation with L-systems. In Proceedings of the 1986
International Computer Music Conference, p. 455-457.
PRUSINKIEWICZ,
P., LINDENMAYER, A. 1990. The algorithmic beauty of plants. New York :
Springer Verlag, 1990.
PRUSINKIEWICZ,
P., JAMES, M., MĔCH, R. 1994. Synthetic topiary. In SIGGRAPH ’94:
Proceedings of the 21th annual conference on Computer graphics and interactive
techniques, 1994, p. 351-358.
PRUSINKIEWICZ,
P., HAMMEL, M., MĔCH, R. et al. 1995. The artificial life of plants. In
Artificial life for graphics, animation, and virtual reality, SIGGRAPH ’95
Course Notes [online], 1995, vol. 7, p. 1-1-1-38. [cit. 2007-04-12]. Dostupné na
internete: <http://algorithmicbotany.org/papers/l-sys.sig95.pdf>.
PRUSINKIEWICZ,
P., HAMMEL, M.. 1997. Visual models of plant development. In ROZENBERG, G.,
SALOMAA, A. (eds.) Handbook of formal languages, vol. 3: beyond words.
Springer-Verlag, 1997.
PRUSINKIEWICZ,
P. 2000. Simulation modeling of plants and plant ecosystems. In Commun. ACM, 2000, vol. 43, no. 7, p.
84-93.
PRUSINKIEWICZ,
P., MUENDERMANN, L., KARWOWSKI, R. et al. 2001. The use of positional
information in the modeling of plants. In SIGGRAPH ’01: Proceedings of the
28th annual conference on Computer graphics and interactive techniques,
2001, p. 289-300.
PRUSINKIEWICZ,
P. et al. 2003. L-systems and beyond. In SIGGRAPH 2003 Course Notes [online].
[cit. 2007-04-12]. Dostupné na internete: <http://algorithmicbotany.org/papers/sigcourse.2003.pdf>.
PRUSINKIEWICZ,
P. 2004a. Art and science for life: Designing and growing virtual plants with
L-systems. In DAVIDSON, C., FERNANDEZ, T. (eds.) Nursery Crops: Development,
Evaluation, Production and Use: Proceedings of the XXVI International
Horticultural Congress. Acta Horticulturae, 2004, no. 630, p. 15-28.
PRUSINKIEWICZ,
P. 2004b. Self-similarity in plants: Integrating mathematical and biological
perspectives. In NOVAK, M. (ed.) Thinking in Patterns. Fractals and Related
Phenomena in Nature. Singapore : World Scientific, 2004, p. 103-118.
QUAN, L., TAN,
P., ZENG, G. et al. 2006. Image-based plant modeling. In ACM Trans. Graph., 2006, vol. 25, no. 3, p. 599-604.
REEVES, W.T.,
BLAU, R. 1985. Approximate and probabilistic algorithms for shading and
rendering structured particle systems. In SIGGRAPH ’85: Proceedings of the
12th annual conference on Computer graphics and interactive techniques,
1985, p. 313-322.
RECHE-MARTINEZ,
A., MARTIN, I., DRETTAKIS, G. 2004. Volumetric reconstruction and interactive
rendering of trees from photographs. In ACM
Trans. Graph., 2004, vol. 23, no.
3, p. 720-727.
Remphrey [online]
[cit. 2007-04-12]. Dostupné na internete:
<http://home.cc.umanitoba.ca/~remphre/research.shtml>.
RUZON, M., TOMASI, C. 2000. Alpha estimation in natural images. In Proceedings
IEEE Conference and Computer Vision and Pattern Recognition, 2000, p.
597-604.
SAKAGUCHI, T.,
OHYA, J. 1999. Modeling and animation of botanical trees for interactive
virtual enviroments. In VRST '99:
Proceedings of the ACM symposium on Virtual reality software and technology,
1999, p. 139-146.
SHEBELL, M.
1986. Modeling branching plants using
attribute L-systems. Master’s thesis. Worcester Polytecnic Institute, 1986.
Zroj: PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic beauty of
plants. New York : Springer Verlag, 1990.
SHLYAKHTER,
I., ROZENOER, M., DORSEY, J. et al. 2001. Reconstructing 3D tree models from
instrumented photographs. IEEE Comput. Graph. Appl., 2001, vol. 21, no. 3,
p. 53-61.
SMITH, A.R.
1984. Plants, fractals and formal languages. In SIGGRAPH ’84: Proceedings of
the 11th annual conference on Computer graphics and interactive techniques,
1984, p. 1-10.
Species XML
File Format [online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.vterrain.org/Implementation/Formats/species.html>.
TOBLER, R.F.,
MAIERHOFER, S., WILKIE, A. 2002. Mesh-based parametrized L-systems and
generalized subdivision for generating complex geometry. In International Journal of Shape Modeling,
2002, vol. 8, no. 2, p. 173-191. Zdroj: BOUDON,
F., MEYER, A., GODIN, Ch. 2006. A survey of computer representations of trees
for realistic and efficient rendering. Report de recherche. In LIRIS, 2006.
TOMAŠKO, I.
2004. Historické parky a okrasné záhrady
na Slovensku. Bratislava : Veda, 2004. ISBN 80-224-0797-6.
trueSpace
[online] [cit. 2007-04-12]. Dostupné na internete:
<http://www.caligari.com>.
ULAM, S. 1966.
Patterns of growth of figures : Mathematical aspects. In KEPES, G. (ed.) Module,
Proportion, Symmetry, Rhythm. New York : Braziller, 1966, p. 64-74. Zroj:
PRUSINKIEWICZ, P., LINDENMAYER, A. The algorithmic beauty of plants.
New York : Springer Verlag, 1990
VAN OYSTAEYEN,
F., GIELIS, J., CEULEMANS, R. 1996. Mathematical aspects of plant modeling. In Scripta botanica Belgica, 1996, no. 13,
p. 7-27. Zdroj: GIELIS, J.
A generic geometric transformation that unifies a wide range of
natural and abstract shapes. In American
Journal of Botany, 2003, 90 (3), p. 333-338.
VĚTVIČKA,
V., MATOUŠOVÁ, V. 1992. Stromy a kry. Bratislava : Príroda, 1992.
ŽÁRA, J.,
BENEŠ, B., SOCHOR, J. et al. 2004. Moderní počítačová grafika.
Brno : Computer Press, 2004. ISBN 80-251-0454-0.
WEBER, J.,
PENN, J. 1995. Creation and rendering of realistic trees. In SIGGRAPH ’95:
Proceedings of the 22th annual conference on Computer graphics and interactive
techniques, 1995, p. 119-128.
Spracované
podľa:
SMOLEŇOVÁ,
K. 2007. Procedurálne modelovanie stromov. Práca k dizertačnej skúške.
Bratislava, 2007. 72 p.